基因的智慧策略
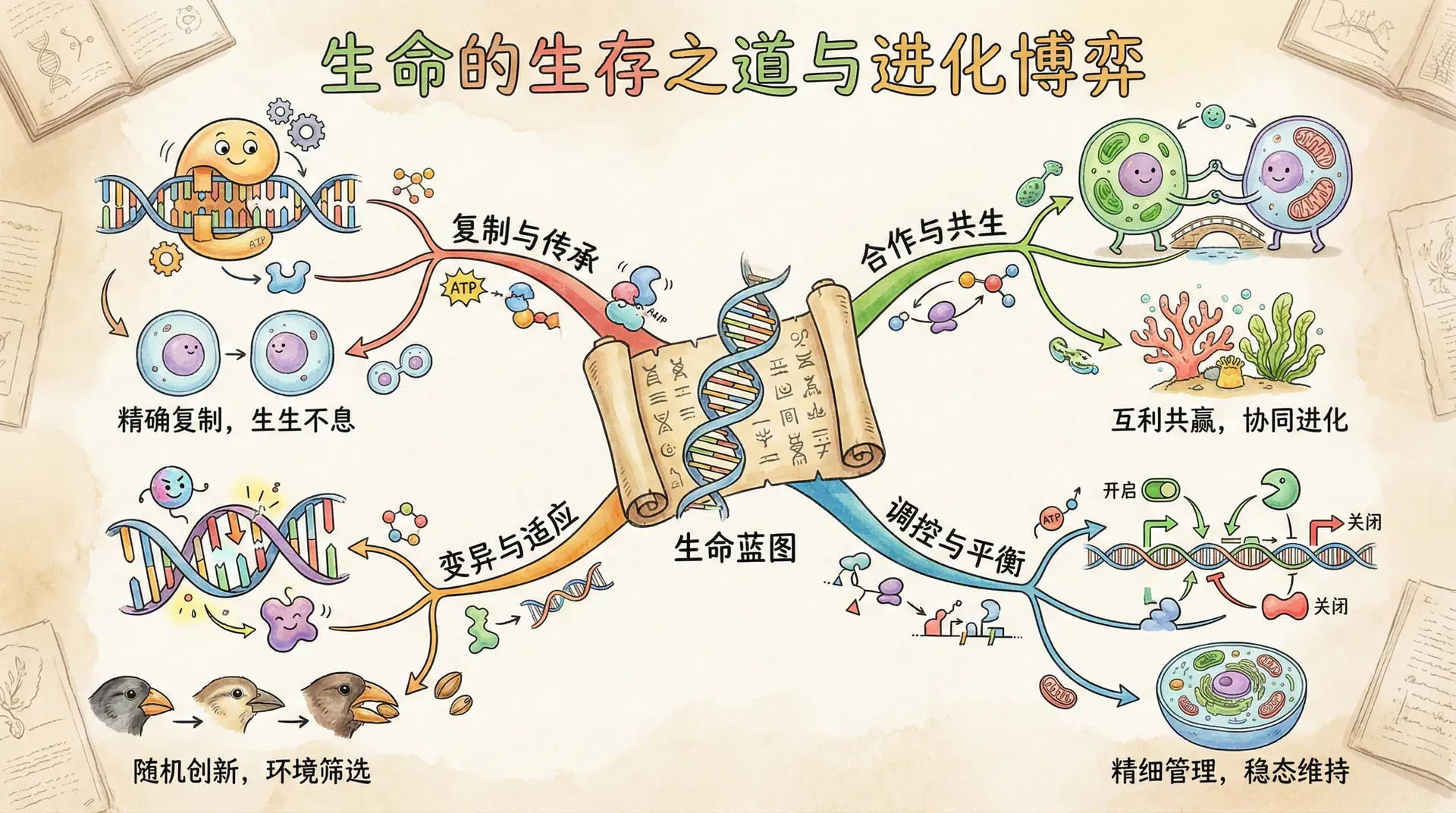
在探讨生物行为的奥秘时,我们首先需要明确一个非常关键的概念:基因并不是某个生物体内一小段孤立的DNA序列,而是在全世界范围内、所有生物个体内、以无数复制品形式存在的“信息单元”。换句话说,你体内的某个基因,很可能也是地球上千万只熊猫、成千上万只老鼠、甚至周围许多人的共同财富。自然选择机制下,真正“努力生存下去”的,其实是基因本身,而不仅仅是个体。
如果我们把基因类比为一个有意识的“策略规划者”(虽然实际上基因没有主观意识,这只是科学上的一种思维工具),那么每个基因都在想方设法让自己在这个庞大的基因库中变得越来越普遍。个体只是基因传播的“载体”和“临时居所”,而基因才是穿越时间与世代的永恒选手。
基因通过“编写”或“控制”其所寄居的生物体行为,来实现生存和繁殖的目标。这意味着,一个基因并不只关心寄主个体是否成功,它还会考虑其他拥有自己复制品的个体。比如说,兄弟姐妹、子女、远亲甚至不相识的同类——只要他们有自己这份基因的拷贝,基因都可能“激励”相关利他行为的发生。
基因识别的策略
在生物进化的过程中,基因需要“辨认”自己的复制品,以便通过促进互助行为提高自身存续几率。科学家发现,基因主要通过两种策略来进行这种识别:一是“标识型”(如绿胡子效应),二是“亲缘识别型”。
绿胡子效应
所谓“绿胡子效应”,是指某一基因通过赋予携带者某种明显的表型特征(如绿色胡子、特殊气味或独特行为),并使携带同样特征的个体彼此间展现出特别的关照或合作倾向。
如果一类老鼠体内存在所谓的“绿胡子基因”,那么这种基因不仅会让老鼠长出非常显眼的绿色胡须作为标识,还会促使这些携带基因的老鼠在遇到同样拥有绿色胡须的同伴时,表现出优先帮助和关照的行为。
现实中,满足这两大条件的“绿胡子基因”极为罕见。原因在于,大多数时候“标识”与“利他激励”来源于不同基因片段,而且表型与行为不易始终一致。进化过程中,这种机制也容易被“作弊者”利用:比如部分体表现出绿色胡须,但并不执行互助行为。
某些真菌和蚂蚁出现过类似效应。例如,红蚂蚁中部分基因让个体仅攻击没有携带该基因的入侵者,从而保护自身基因。尽管如此,这种机制远不如亲缘识别广泛和有效。
亲缘关系识别
在自然界,更加普遍和稳健的方式是通过血缘关系来“识别”自己的基因复制体。简单来说,血缘越近,共享基因的比例越高,互助行为也更容易发生。哺乳动物、鸟类甚至昆虫等,都表现出基于亲缘关系的照顾或牺牲行为。
常见的亲缘互助行为:
- 老虎会不顾危险保护自己的幼崽
- 松鼠在遭遇天敌时会用高声警报提醒亲属
- 蜜蜂工蜂为蜂后与同巢兄弟姐妹奉献生命
下表展示部分生物在亲缘识别下常见的行为与对象:
研究发现,这些亲缘行为并不仅仅是文化或习惯产物,而是深植于基因策略之中的本能。它也帮助我们理解为何人类社会中的家庭、宗族纽带如此紧密——基因倾向于鼓励亲属间互帮互助,从而提升自身在群体中的扩散概率。
我们不仅在动物世界看到这些现象,事实上许多人性中的亲情纽带、家庭归属感,也都带有这样深刻的生物学根基。
亲缘关系的数学计算
基本概率原理
要理解亲缘关系中的数学本质,我们先从一个简单案例入手。假设在一个典型的中国家庭中,你携带了一个非常罕见的基因G,那么这个基因必然是从你的父亲或者母亲遗传而来,每个方向的概率都是50%。人类每个子代只从双亲中各获得一半基因材料,而每个父母本身也只将自身基因的一半传递给每个子女。
假设基因G来自你的父亲。你父亲产生的每一个精子,有一半带有基因G,一半不带。因此,你的亲兄弟姐妹出生时,同样有50%概率获得这个基因。这一推导法则适用于任何遗传位点,也适用于母亲方向。进一步来说,你和每个亲兄弟姐妹之间,平均有一半的基因是共享的。
把这种思路推广到祖父母、叔伯、表兄等更远的亲属,我们其实可以一直追溯到最初的共同祖先,计算每一代遗传时“掷硬币”的概率累积,从而得到精确的血缘概率。
亲缘关系指数
在遗传学中,我们用一个“亲缘关系指数”(coefficient of relatedness, 通常记为r)来衡量两个人共享同一基因的概率。这个值越高,说明两者之间基因共享越多。下面这个表格展示了常见亲缘关系的指数与共享基因概率:
例如,你和同卵双胞胎的基因理论上是完全一致的,r=1。而与亲生父母或亲兄弟姐妹,则有一半的基因相同,r=0.5。再远一步,与祖父母、孙辈,或是叔伯、侄子女则是0.25,表兄表妹之间则只有0.125。
计算方法
那么,如何具体计算两个个体(比如A和B)之间的亲缘指数呢?可以按照如下步骤:
- 找到共同祖先:首先确定A和B最近一次共享的祖先,比如亲兄弟姐妹的共同祖先是父母,表兄弟的共同祖先则是祖父母。
- 计算世代距离:从A出发,经过各自的祖先到达最近的共同祖先,再从该祖先降到B。这个路径的“步数”就是总的遗传路径长度。例如,表兄弟之间是A→父母→祖父母→B的父母→B,一共4步。
- 应用概率公式:每一步遗传都乘以 。总的亲缘关系指数就是所有路径上的概率相加,每条路径的概率为 。例如,父母-子女之间是 ;叔伯-侄子女的路径为2步,。
举例:
- 直接亲兄弟姐妹:A(你)父母B(兄弟姐妹),需两步,每个路径概率 ,但由于有两个父母,每条都算一条路径,合起来 。
- 表兄弟姐妹:需要通过父母到祖父母再到堂表兄,全程4步,概率 ,但可有多条路径,最终加总为 (12.5%)。
这种计算法则可广泛应用于真实家族树,帮助科学家精确评估基因传播效应和行为动机。
野生动物中的亲缘选择
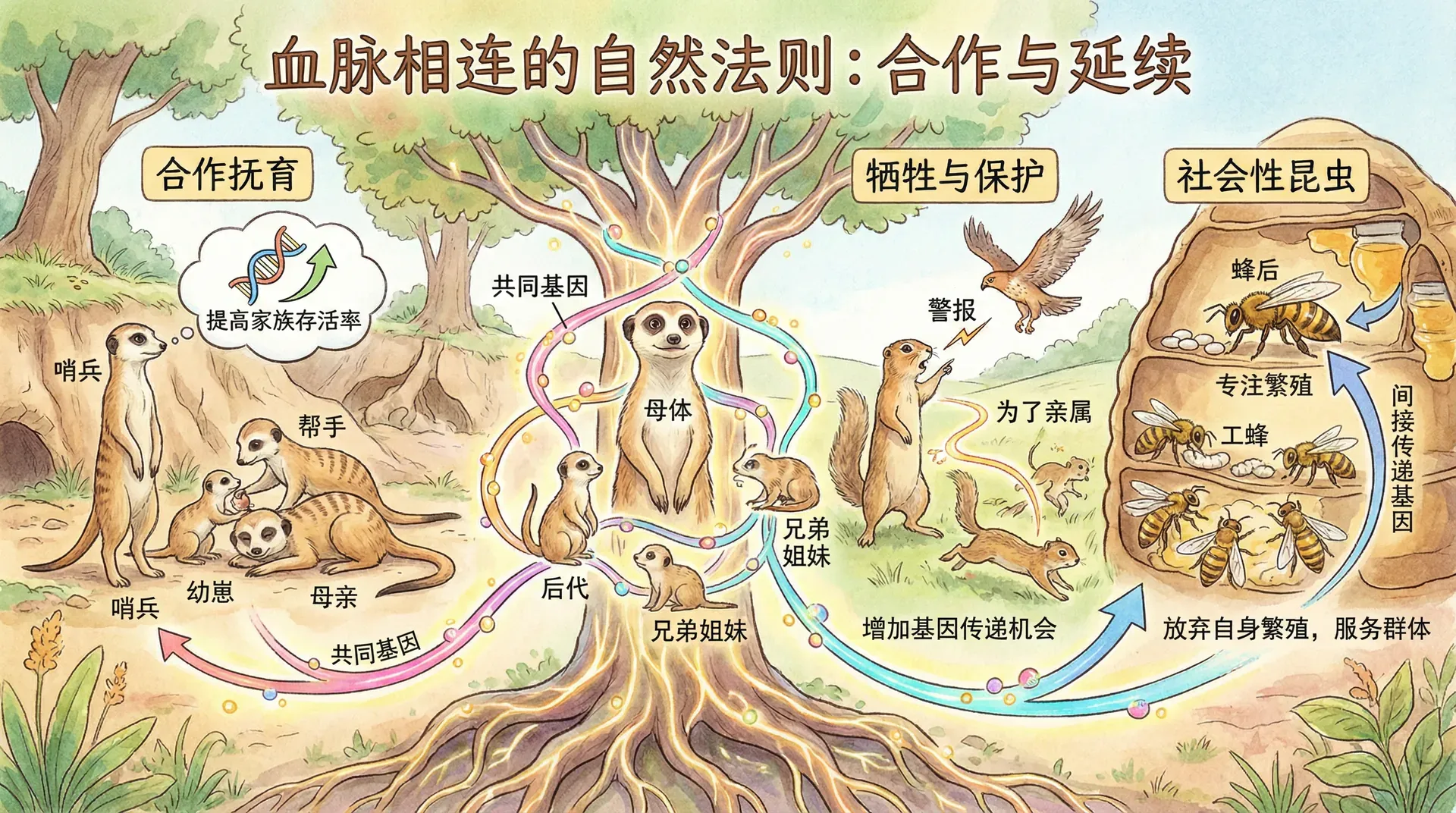
亲缘选择并不仅仅是一种理论假设,而是在野生动物世界中有大量实际案例支撑的进化机制。下面我们通过具体的生物实例更深入地理解亲缘选择的力量。
川金丝猴的群体行为
在四川的自然保护区中,研究人员长期跟踪观察野生川金丝猴,发现了极具代表性的群体利他行为。当豹子等天敌出现时,成年雄性金丝猴常常会主动发出警告叫声,并选择留下来吸引掠食者注意力,而雌性及幼崽则趁机逃离。这样的行为表面上是“牺牲自我保护他人”,实际上体现了深刻的亲缘选择逻辑。
为什么会这样?研究者通过基因分析发现,一个成年雄性与同群的其他成员平均亲缘关系大约是0.3——也就是说,每救下一个群体成员,自己基因的间接收益就是自身的30%。如果它能保护到多个亲属,综合的基因“回报”就超过了自己的风险或损失。这种舍己救亲的行为不仅见于金丝猴,在诸如狒狒、狼、象等其他群居哺乳动物中也都有类似现象。
扩展来看,群体规模、血缘密度、以及天敌威胁等因素,都会影响亲缘选择带来的协作强度。有些学者还发现,群体内亲属比例越高,警戒与协作行为越频繁;反之,群体成员的亲缘关系越疏远,个体则更趋于自保。
中华白海豚的救助行为
在珠江口广阔的水域,中华白海豚形成了相对封闭的种群。研究者多次目击这样的场景:当一只海豚受伤、虚弱到无法自主浮出水面时,其他健康的海豚便会有序地围绕并托举伤者,将它抬至水面呼吸。整个过程可能持续数十分钟到数小时,期间托举的责任会在亲属间轮流转换,显示出高度协作性。
这种现象极好地展现了亲缘选择理论的应用。由于中华白海豚的近亲繁殖程度高,群体内成员之间共享大量基因。对单个个体的协助实际上是在保护整个家族、甚至整个基因库的存续。因此从“基因的自私”角度来看,这种群体救助是高度“理性”和进化上有利的选择。
类似的亲属救助还见于海豚之外的其他社会性动物,比如大象群体间的帮助、非洲野犬之间的照顾等,显示出亲缘选择原理在自然界的普遍性和多样性。
识别亲属的实际方法
但动物并不会像人类那样进行家谱统计。那么它们靠什么规则辨认彼此的亲缘关系?研究发现,大多数物种都会运用一些简单但有效的“经验法则”进行亲属识别:
有些物种甚至发展出了复杂的“家庭代码”,如部分鱼类或昆虫能区别出卵块/蚁巢中的自家与外家成员。亲属识别虽非绝对精确,却已足以让进化选择“押注”亲缘带来的行为红利。
欺骗与反欺骗
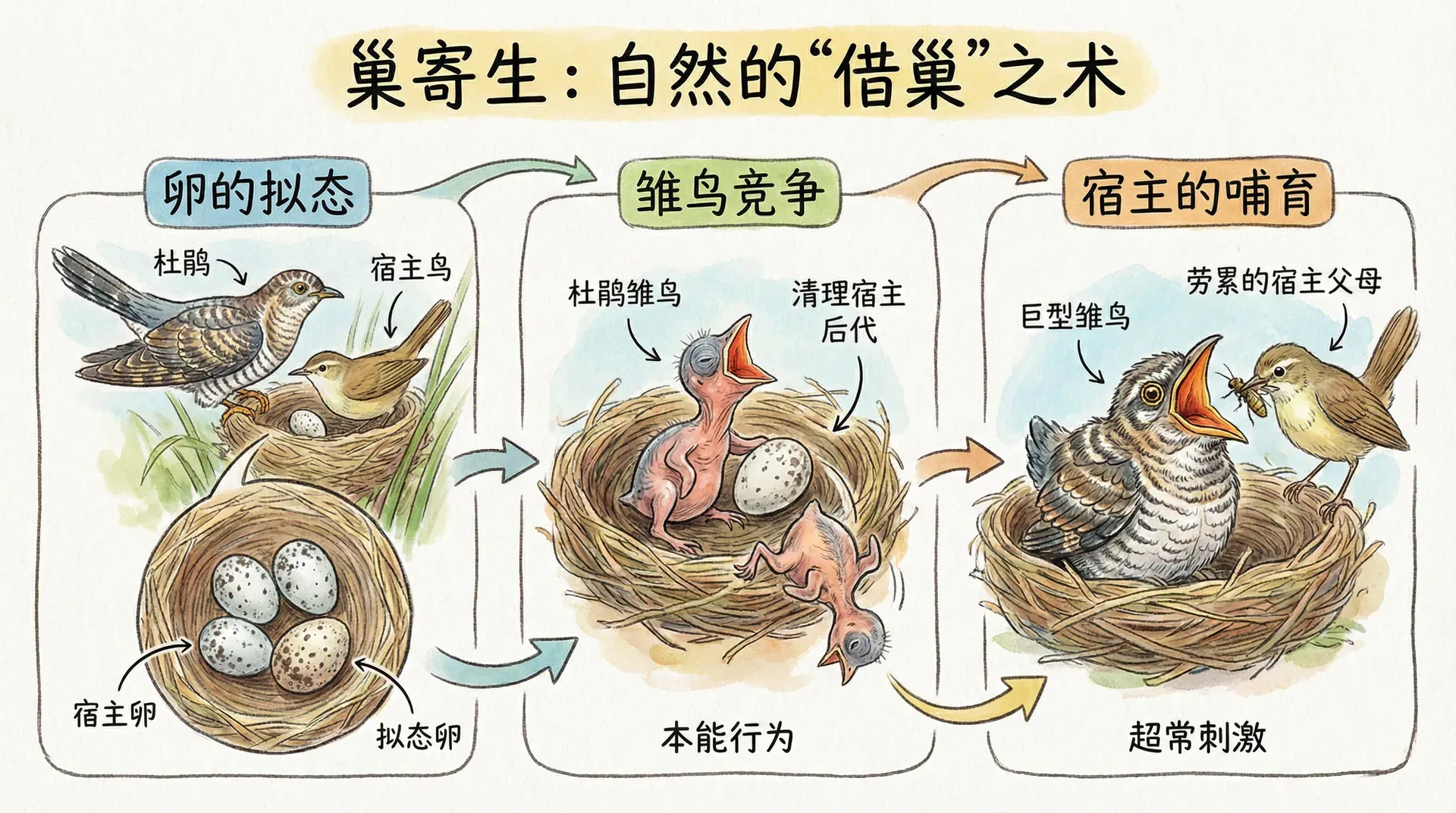
杜鹃鸟的寄生策略
并不是所有动物都会自觉地照顾亲属。自然界中还存在对亲缘系统加以“利用”的欺骗现象。例如,在中国及东亚地区广泛分布的大杜鹃,演化出一种著名的“巢寄生策略”:雌性大杜鹃不自己抚育后代,而是专门将蛋产在麻雀、伯劳等其他鸟类的巢中,让“受骗”的宿主负责孵化和喂养。
大杜鹃的蛋不仅形状、颜色、斑纹极度模仿宿主的蛋,有时甚至能模仿出光泽和微小杂点。这一切都是为迷惑宿主,使其无法分辨出“冒牌货”。受害的宿主鸟类基于对“自家雏鸟”的本能养护,会用辛苦采集来的食物养育杜鹃幼仔,尽管后者很可能与自己没有任何亲缘关系。
这个例子展示了演化军备竞赛的过程:
- 大杜鹃的策略:通过自然选择,蛋的外观伪装越来越逼真,甚至雏鸟唤食信号也进化得更像宿主后代;
- 宿主的反击:宿主们进化出更敏感的识别能力(比如记忆自己的蛋数、注意色差),发现可疑“冒牌蛋”时直接将其推出巢外;
- 持续进化:杂交、错杀亲属等进化压力推动双方在识别与伪装能力上螺旋式提升,形成“识骗与被骗”的动态平衡。
这种“欺骗”现象提醒我们,自然界中的利他行为绝不是道德信仰下的无条件付出,而是基于基因利益的权衡。当识别机制受限(如无法区分自家和外来后代)时,亲缘选择策略就可能被“钻空子”——也即是说,生物进化中既有合作,也有无休止的对抗,这正是自然世界的真实样貌。
父母照顾 vs 兄弟姐妹互助
为什么父母照顾更普遍
乍看之下,父母-子女关系和兄弟姐妹关系在遗传学上似乎是“等价”的:无论是亲生父母和孩子,还是全血的兄弟姐妹,平均都共享50%的基因片段(r=0.5)。但令人疑惑的是,在自然界几乎所有已知动物中,父母对后代的照顾行为远比兄弟姐妹之间的无私互助要常见和强烈得多。这是为什么?
1. 确定性更高
父母,尤其是母亲,几乎可以百分之百确定自己的孩子携带了自己的基因。而兄弟姐妹之间,虽然理论上有50%的基因共享,但在某些情况下,这一共享是基于概率统计。例如,在多配偶或群体繁殖的种群里,兄弟姐妹可能并非全血;而母亲对自己所生的后代具有基因关系的高度把控。这种确定性使得自然选择更容易“信任”父母的投入。
2. 能力与资源优势
成年个体(父母)不仅有更强的体力、经验和社会地位,往往也能更有效地找到食物和作战防御。因此,对幼体来说,来自父母的照顾能显著提高生存率。相比之下,幼小的同胞姐妹往往缺乏资源与能力去提供真正意义上的帮助。
3. 时间和繁殖价值
在进化论的“投资回报”计算中,年轻后代比年长个体有更大的预期繁殖潜力。对已经成年且生育期即将结束的父母来说,投资于子女能让自己的基因延续更远。兄弟姐妹之间尽管也能通过互助间接促进基因的保留,但在残酷的自然环境下,“自救”或保护新一代后代优先级更高。
4. 演化的稳定策略
动物行为学的研究表明,父母照顾后代是一种“进化上稳定”的策略(ESS)。如果整个种群中广泛存在父母照顾行为,个体稍作改变就会处于不利地位,比如“懒惰”的父母后代存活率更低,很快被淘汰。
兄弟姐妹互助行为的典型与限制
当然,也有许多动物物种表现出“兄弟姐妹互助”:蜜蜂工蜂会集体照料同窝幼虫,川金丝猴、狒狒等灵长类也会出现年幼成员相互搀扶、警戒乃至共同御敌的现象。但这些行为往往建立在极高亲缘密度的基础上(比如单一蜂后控制整个蜂群,全部工蜂都是同母异父/同母同父的姐妹),或者兄弟姐妹行为是父母行为的功能延伸。
在有些物种(如部分鸟类),年长的兄弟姐妹会协助父母喂养刚破壳的弟妹,有学者称之为“协助繁殖”或“辅助育雏”。但总体来说,兄弟姐妹间的“利他”行为更容易因个体利益冲突、营养分争夺等问题而受限制。甚至在某些鱼类、蛇类里,兄弟姐妹之间还会出现“同胞竞争”甚至“同类相食”。
中国传统文化中的体现与深层意义
中国传统中,孝道理念极为重要:子女对父母的赡养和尊敬是社会道德的核心。这乍看与进化生物学的“父母更应优先投资子女”有所出入。实际上,这说明:人类社会的行为已受到复杂的文化、历史、宗教影响,纯粹以基因为中心的解释只能揭示部分底层驱动力。
但如果观察各地风俗,仍可发现一些符合亲缘选择理论的现象——如家族内部(亲兄弟姐妹之间)互助的一致性、家产继承优先考虑直系后代等等。这些举措本质上有助于最大化家族基因的留存率,也就是生物学上“亲缘选择”的社会化表现。
父母对子女的投入如此深厚,从怀胎、哺乳到长期陪伴,非常符合进化上对于基因延续的“最佳投资”。而兄弟姐妹之间的帮助,则需在利益权衡和资源斗争中找到平衡点——动物行为学、心理学、人类学都在这个问题上有更多值得探究的细节。
实际应用与启发
对野生动物保护的启示
理解亲缘选择机制,对野生动物保护具体策略具有极大现实意义:
许多野外研究(如圈养与野化交替、断代放归等)都证实,亲缘结构完备的群体,更容易展现防御天敌与合作养育等进化优势,对种群复壮和遗传多样性维持有重要作用。
对人类社会行为的理解和借鉴
亲缘选择理论不仅帮助我们理解动物,也为现代社会中人与人之间的合作、信任与冲突提供了底层解释:
- 为什么亲属间互帮互助更容易发生?家族企业为什么往往基业长青?这是基因与亲缘信任在现实中的投射。
- 血缘纽带为何被广泛用于塑造认同感、归属感乃至社会组织结构?这是千百年进化过程中“利他基因”潜在作用的社会版本。
- 人类社会逐步发展出“不以亲疏论好坏”的道德规范(如公平、公益、契约),正是在超越基因驱动力的基础上构建更加文明化、多元化的社会生态。
理解这些生物学原理,并不意味着我们要把所有人类社会行为都“归咎于基因”——而是帮助我们在追问“为什么如此”时,拥有一个更深的解释层面,从而实现理性与情感、传统与现代的更好融合。
总结
亲缘选择理论为我们描绘了生物世界中利他与合作的核心动力,从川金丝猴奋不顾身救群,到中华白海豚温柔托举同伴,从杜鹃鸟的寄生欺骗到大熊猫母性的付出,无一不印证着“基因自私”下的进化智慧。
更重要的是,这个理论不仅解释了自然界中的动物行为,也做出了重要的人类社会启示。它提醒我们:文化、道德与教育让人类超越了单纯基因本能,我们可以因选择、因制度而开创新的社会合作模式。基因与文化,共同塑造了我们的今天。
亲缘选择既展现了动物界利他行为的本质,也提醒我们理解自己与他人的深刻联系——每一个生命个体,都是无数祖先基因竞合的产物,也承担着未来世代的希望和责任。