生存竞争中的策略选择
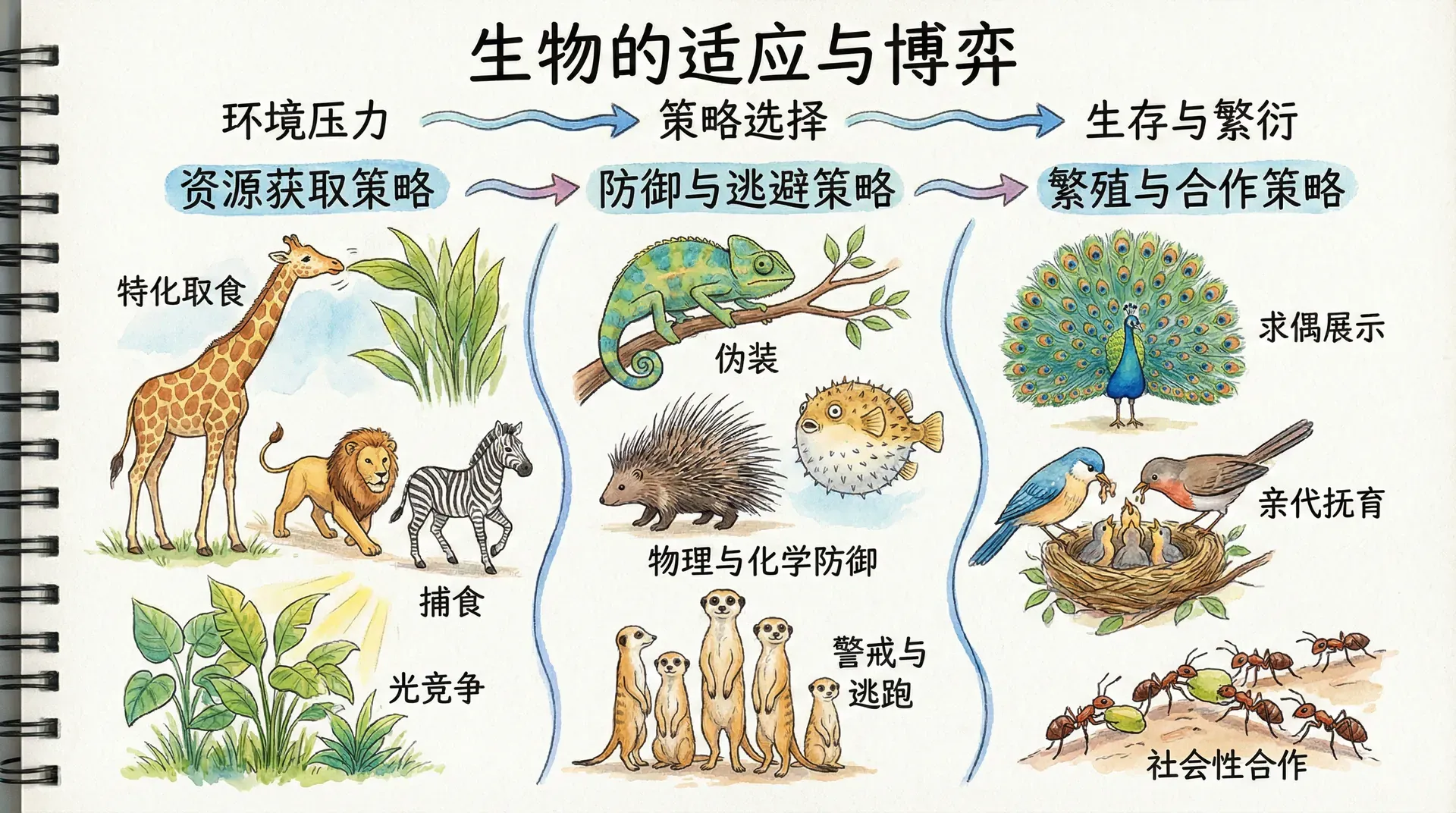
在自然界中,生物个体之间的竞争几乎无时无刻不在发生。从陆地到天空,从森林到海洋,每一个生物体为了生存与繁衍,都不得不与同类甚至异类之间展开各种形式的争夺。比如,一只麻雀需要和其他麻雀争夺食物、栖息地,还要防备天敌;一头狼即使身处自己的族群中,也难免要与其他成员竞争配偶或争夺带头者的位置。而这一切表面看来就像“弱肉强食”的简单规则,实际上背后却隐藏着极为复杂的生存策略和博弈。
自然界的竞争不仅仅体现在动物之间,植物也在进行着激烈的较量。比如树木为争夺阳光向上生长,藤蔓植物则会缠绕、遮蔽邻近的植株。此外,某些昆虫会通过信息素影响同类的行为,细菌也可能通过分泌毒素抑制其他菌群扩张。由此可见,“竞争”是整个生命世界中普遍和基本的现象。
为什么不是殊死搏斗
当我们观察动物之间的争斗时,会发现一个耐人寻味的现象:大多数情况下,动物并不会拼个你死我活。譬如两只公鹿在森林里为了争夺配偶,往往只是角对角顶撞、展示气势,极少真的将对方置于死地。又如鸟类争食时,很多时候只是通过叫声、亮羽和威吓动作一决高下,罕见真正致命的冲突。这究竟是为什么?
理论上,从“自私基因”的视角来看,个体似乎应当千方百计消灭一切潜在的竞争对手,务求将自己的基因最大化地传递下去。可是在真实的自然界里,这种“赶尽杀绝”的策略并不普遍。其根本原因在于,激烈的冲突会带来巨大的成本,包括受伤、死亡、精力消耗甚至被天敌趁虚而入。与此同时,完全的胜利并不一定意味着生存优势,有时候反而会因消耗过度而落于下风。
假设在一个池塘边,三只野鸭A、B、C都想占据最佳的觅食位置。如果A选择攻击B,并且成功将B赶走,表面上A得到了短期的利益。但其实,旁边的C才可能是最大的赢家——因为竞争对手减少了,自己反而能轻松获得更多资源;而A则因战斗消耗掉体力,还可能受伤,得不偿失。类似的策略博弈在自然界屡见不鲜,这种复杂的生态关系也促使了各种非致命竞争行为的演化。
实际上,许多动物在争斗前后都会进行复杂的试探和威胁展示——从鲤鱼的腾跳、鸟类的鸣叫,到哺乳动物咆哮、立毛,这些都属于非致命的“武力展示”。它们的出现正是因为死斗的代价太大,而自然选择促使个体倾向于选择成本更低、风险更小的解决办法。
在复杂的竞争关系中,消灭一个对手可能让其他对手获益更多,而未必让自己获得最大的利益。因此,很多时候动物更倾向于采取有限度的竞争、展示和威慑,而不是直接拼至你死我活。
斗争的成本与收益
让我们以两只雄性东北虎争夺领地为例。这片领地资源丰富,充满了猎物和水源,能够极大提升占有者的生存和繁殖机会。那么,面对竞争对手,一只老虎会怎么决定是否要全力以赴地发起攻击呢?背后的考量其实远比直观想象复杂得多。
-
即便激烈进攻最终赢得了领地,也存在许多潜在的不利后果。例如,如果它在打斗中严重受伤,捕猎能力会大幅下降,长期甚至可能因无法捕食而饿死。更轻的伤势也会影响行动速度或警觉性,增加被其他掠食者或竞争对手袭击的风险。此外,受伤的老虎还可能因为虚弱失去对领地的掌控,最终落得“为他人做嫁衣裳”的下场。
-
战斗过程不仅消耗巨大的体力和能量,还会占用大量时间。这些时间本可以用来觅食、巡逻或休息,甚至进行交配和照看后代。而体力消耗过多,也可能使老虎在后续的竞争与生存中处于劣势。与此同时,长时间的冲突和嘶吼也可能吸引天敌或其它潜在对手的注意,无形之中增加了外部威胁。
-
面对对手时还必须纳入对手实力的评估。要知道,一只能够长期占据优质领地的雄性老虎,往往本身就拥有极强的实力和丰富的战斗经验。贸然挑战可能意味着面对一位久经沙场的强敌,失败的风险和损失也随之翻倍。
除了上述直接后果,还有一些间接但同样重要的“隐性成本”。比如,打斗的噪音和气味可能干扰群体的稳定,影响附近雌性的择偶行为,进而间接影响自己的繁殖成功率。有时候,即使自己取胜并赶走对手,剩下的第三方竞争者反而可能趁机渔翁得利。
因此,是否发起战斗需要在大脑中瞬间进行一番“精密计算”:
可以看出,动物在每一次冲突前都要权衡成本与收益,作出理性的选择。这种“精明计算”并非真的需要复杂的思考,而是基因在代际演化中塑造出的行为趋向。最终,这种动态权衡过程促使动物发展出了各种复杂、灵活的竞争策略:有时选择直接对抗,有时以威吓展示结束冲突,有时干脆选择回避与等待更好的时机。
在个体与环境、对手以及局势的不断变化中,不同的选择导致了自然界丰富多彩的行为和社会结构。这也正是生物多样性和生态平衡的基础之一。
进化稳定策略
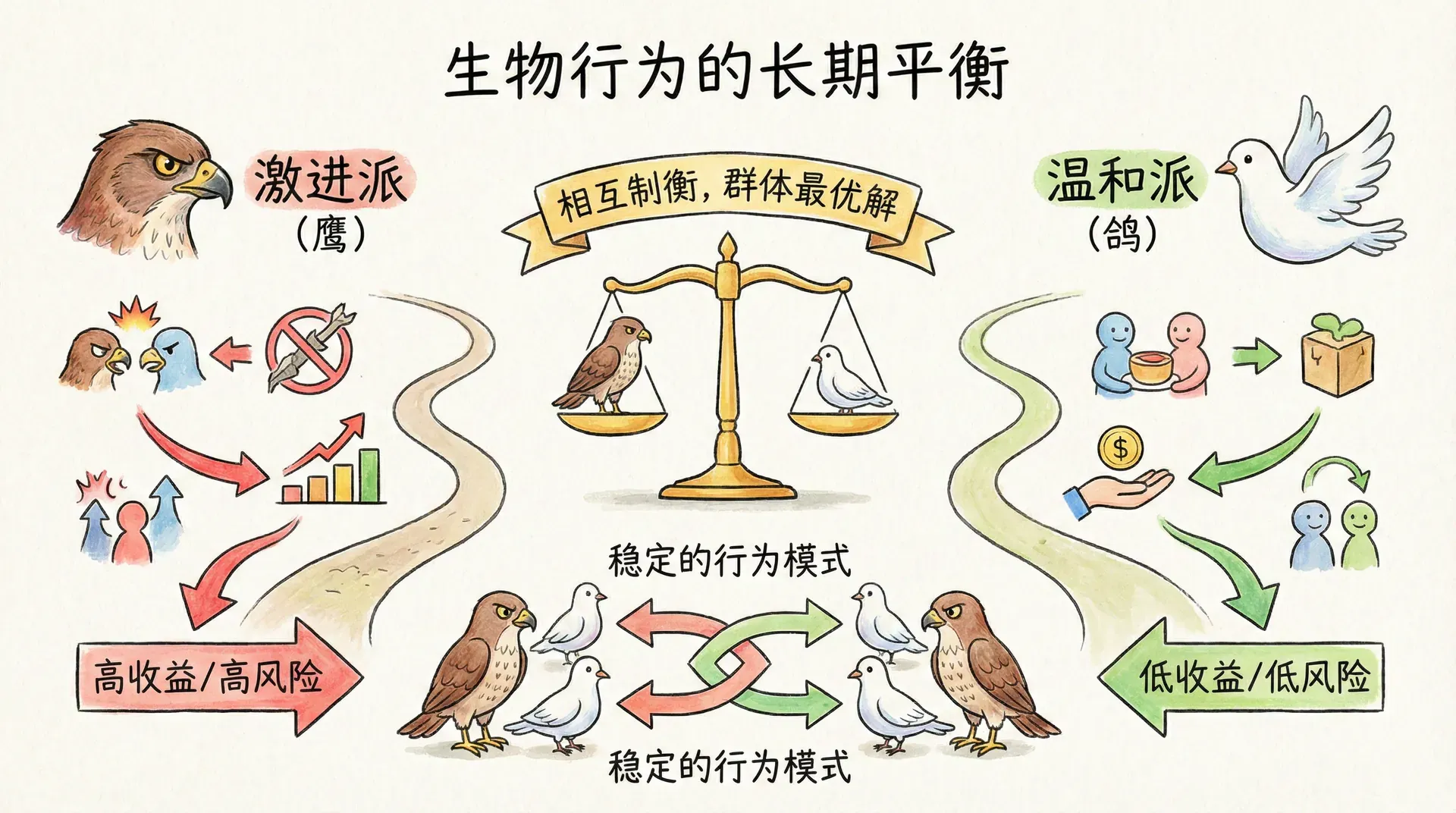
英国生物学家约翰·梅纳德·史密斯(John Maynard Smith)在1970年代提出了“进化稳定策略”(ESS,Evolutionarily Stable Strategy)的理念,为我们理解自然界中动物行为的演化提供了强有力的分析工具。ESS指的是这样一种策略:如果几乎所有个体都采用它,任何新出现的替代策略都无法通过“入侵”获得更高的进化优势、最终取而代之。换句话说,ESS是一种抵御异类突变和行为扰动的行为模式,它解释了为什么许多看似复杂甚至带有“妥协”色彩的行为能长期稳定地存在于自然界。
用模型来理解
虽然这个概念有点抽象,我们可以用“鹰-鸽博弈”模型来说明。假设一个群体中,个体间为了争夺资源(比如食物、领地或配偶)进行竞争时,只存在两种典型行为:
鹰派(Hawk):遇到竞争,就发动全力攻击,不达目的不罢休,除非自己受重伤或者对方逃跑。
鸽派(Dove):遇到竞争时,只做威胁展示,不会真的攻击,一旦对手选择升级冲突,自己就会退让。
我们为各种对抗结果分配“分数”,这些分数代表遗传适应度、后代数量的期望变化或者长期资源收益:
全是鸽派的世界
让我们从极端情况开始思考:假设群体最初全是鸽派。当两只鸽派遭遇时,它们通过持久的威胁舞蹈比“气势”,但大家都避免真正开打。获胜方拿到资源(+50分),但要付出时间成本(-10分),共得40分;失败方则只是失去10分时间。鸽派之间的平均收益=(40+(-10))/2=15分。
但如果此时群体里出现一只“鹰派”突变体会怎样?每当它遇到鸽派,就直接抢走资源,鸽派立刻退让不受伤,鹰派轻松获得+50分。显然,新的鹰派基因有极大优势,迅速扩散至群体。
全是鹰派的世界
随着鹰派增加,假如所有个体最终都变为鹰派。结果会变得惨烈:鹰派遇到鹰派,需要通过流血战斗分胜负。胜者得+50分,败者-100分,取平均=(50 + (-100))/2 = -25分。每次冲突,平均下来所有鹰派成员都付出负收益,整体适应度下降,生存风险反而增大。
此时,如果再有鸽派突变体出现,他选择威胁一下看到对方很猛就退让,不会受伤,每次得分虽然是0,但却远高于鹰派-25分。所以鸽派策略又会被选出来。如此来回循环,单一策略都难以独占优势。
数理上的“稳定平衡”
通过详细的数学分析,我们可以算出,只有两种策略(鹰、鸽)在特定比例下共存才是“进化稳定”的。在上述设定下,稳定点的近似比例为7:5,也就是在进化的长期博弈后,大约58%的群体成员采取鹰派,42%采取鸽派。在这个比例下,两种策略的平均收益刚好相等,谁也无法轻易取代谁。
上图展示了随着鹰派比例的增加,鹰派和鸽派各自的平均收益变化趋势。交点恰好是进化稳定点。这个点并不意味着“群体整体最好”,而是任何试图“入侵”的新策略都难以取代现存格局。
进化稳定策略强调“难以被打破的平衡”,而不是所谓“对大多数个体最友好”或“整体适合群体利益”的方案。这意味着,某种策略是否ESS,只和自身抵御其他策略的稳定性有关,不必顾及“整体最大化”。
更复杂的策略
当然,现实中的动物行为远非只有鹰和鸽那样简单。自然界还存在许多条件性、混合性和更为精细化的策略。以下是常见的几种变体:
(1)报复者(Retaliator):初始采取鸽派模式,只做威胁展示,若对方攻击,自己则变为鹰派反击。这样既可避免无谓冲突,又能保护自己不被敌人占尽便宜。
(2)欺软怕硬者(Bully):平时像鹰派那样咄咄逼人,但一旦遇到对手还击,自己反而马上撤退。
(3)试探者(Prober):一开始像报复者,但偶尔主动“试探”升级冲突,观察对方反应;如果对方坚决反抗则收敛,否则持续进攻。
除了这些,理论和模拟研究还发现,如“随机混合使用两种策略”、“根据上次交锋结果调整行为”等复杂机制都可以形成进化稳定结构。
当这些策略在个体之间自由演化、相互竞争时,理论和大量计算机模拟都显示:报复者策略最为稳定。如果一个群体几乎全是报复者,其他策略很难入侵或立足。这与自然界中常见的现象吻合:绝大多数动物的攻击都带有“节制”,威胁多于真打,只有在被真正攻击时才会坚决反击,从而避免高风险、零和的消耗。
进化稳定策略的思想还为我们理解动物为何普遍存在“威胁行为”、“仪式化冲突”与“有限打斗”等现象提供了理论依据。强烈的致命对抗稳定概率低,适度威慑加上报复性反应最为广泛。
领地行为的本质
在自然界中,动物“守护疆土”极为常见。这不仅关乎资源的占有,更隐藏着深刻的博弈机制。例如:
事实上,如果鸽子、麻雀等其他物种进入画眉鸟的领地,画眉鸟往往不会表现出激烈的反应;真正激烈的斗争通常只发生在同种个体之间。这种选择性防御背后,体现的是利益最大化的行为进化和减少冲突的策略智慧。
著名动物行为学家尼古拉斯·廷伯根(Niko Tinbergen)曾做过一个经典实验,揭示了“领地优势”的原则。他把两只雄性刺鱼分别安置于水族箱的两端,各自“安家筑巢”。随后,将它们分别放入透明试管,再把试管并列移动。实验发现:
- 试管靠近“鱼A的领地”时,鱼A会主动作出攻击姿态,鱼B则表现出退缩,即使两鱼被隔离在试管内。
- 反向靠近“鱼B的领地”时,行为立刻调换。
实验展示了“先到者优势”(bourgeois strategy):谁在自己的主场谁就选择攻击,外来者则选择撤退。这一策略避免了反复和高风险的争夺,形成了不起争议的分辨规则,如下所示:
由于“先来后到”带来的不对称性,动物们能借助“场所归属感”快速做出决策,常常在威胁展示阶段就化解了冲突,极大减少了资源和精力消耗。这一机制适用于鸟类、哺乳动物、昆虫等多种生物。例如,斑马鱼、燕雀、鼩鼱等在无视觉标记的情况下,也会在熟悉区域表现出更强的攻击性,而陌生来客会主动撤离。
领地行为的核心并非“领地本身极其有价值”,而在于先后顺序带来的博弈利益不对称。正因为有了省力的规则,动物们无需反复激烈拼杀。
体型差异的影响
现实中的动物个体,体型和力量往往具有显著差异。这直接决定它们在遭遇时的策略选择。观察发现,多数冲突其实在“气势”展示阶段早已定胜负。比如两头雄性麋鹿对峙时,一方如明显高大健壮,另一方干脆主动退让,仅在旗鼓相当时才陷入激烈缠斗。
动物本能倾向“实力判别+避免受伤”策略,它们会通过视觉(展示体躯)、鸣叫、动作等“信号”评估对方实力。如果判断自己劣势,就会及时撤退,以避免不必要的受伤与日后繁殖机会的丧失。下表展示了不同情况下合理的策略选择:
此外,学界曾提出“反常策略”作为脑力实验——如果一个群体中个体都只攻击比自己强(而非弱)的对手,结果反而是没有个体真正会受伤,因为强者会自动躲避、弱者攻击却无法真正获胜。表面上看每个人都很“进攻”,实际上没有“战斗”发生。然而,只要有一个“正常策略”者出现(遇弱进攻、遇强回避),这种反常格局会被击破,反常者立即陷入伤害,不再具备稳定性。
在自然界中,不同动物在发生冲突前,普遍会通过一系列信号和仪式来“评估”彼此实力,有效避免了真正的激烈搏斗。例如,大猩猩在正式打斗前,通常会通过捶胸等夸张的动作展示力量,从而让对方判断是否值得一战,这极大地降低了生死对抗的概率。同样,公鸡在决斗前也会通过竖起羽毛、绕圈行走和鸣叫等一整套仪式化行为展示气势,双方据此评判形势,只有旗鼓相当时才可能爆发真正的争斗,这种“显摆”大幅降低了因冲突导致严重受伤的风险。甚至在鱼类中也可以发现类似机制,如七彩神仙鱼可通过体色的变化和鳞片的光泽来传递自身实力,进而影响对方的行为选择。这些策略让动物能够识别优势与劣势,优先选择撤退或展示,而不是直接陷入危险的搏杀。
此外,体型差异还会因年龄、性别、营养状况动态变化。例如,幼年动物几乎总会让位于成年个体,雄性个体会争夺并划分较为严格的等级和领地。
“遇强则避,遇弱则战”是许多动物群体的进化稳定策略。通过“显摆赛”、“叫声较量”,动物能够高效辨别对手实力,既达成利益最大化,又减少伤亡和能量消耗。
记忆与等级制度
除了体型和力量之外,一些动物还具备记忆和学习的能力,能够记住自己与其他个体交往的历史结果,从而形成群体中的等级秩序。
研究发现,一只经历了多次胜利的蟋蟀会变得越来越好斗,而多次失败后则趋向胆怯。这表明动物可以通过过去交锋的结果调整行为倾向。科学家曾用假蟋蟀反复“打败”实验蟋蟀,之后这只蟋蟀在面对真实竞争者时也变得不敢作战,进一步证实了社会记忆的作用。
这种依赖记忆的机制促成了稳定的社会等级。即使起步时个体差异很小,持续的互动也能拉开地位分化:高排名个体变得更自信,低排名更怯懦。部分群体中还会形成“赢家效应”和“输家效应”,例如赢了一次更容易继续赢,而输了则更易连续失败,社会结构由此逐步固化。
母鸡群体里出现的“啄序”现象,是社会等级的经典案例。当一群不熟悉的母鸡被关在一起,通常会经历短时期的混乱性斗争。熬过初期后,每只母鸡都能认清自己在群体中的“地位”:有权啄其他鸡而不被回击的“老大”,以及只能被啄却无法还手的“老实鸡”。这种复杂的社会记忆极大地减少了群体中反复的暴力,一旦权力排序稳固下来,争斗会显著减少。
这种等级序列实际上并不是为了“群体和谐”而演化出来的,而是出于每一个体都期望最大化自己的生存和繁殖利益。当等级秩序成立后,动物们减少了重复性“无意义内耗”,能够把更多的能量和资源用于成长、生殖和寻找食物。以母鸡为例,有研究发现,在稳定群体中产蛋量显著提升,并不是因为“集体意识”,而是由于减少了能量消耗和受伤风险。
上图展示了两种情形下母鸡群体“打斗次数”的变化。可以看出,在成员固定的群体中,打斗频率迅速下降,极快形成等级体系。而成员频繁更换的群体,权力结构不断被打乱,打斗始终难以消退。稳定的社会结构能够显著减少能量损耗,提高生活效率,这也是等级制度在许多动物社会持续存在的重要原因。
不同物种之间的竞争
同一物种的个体之间的竞争通常是最激烈的,因为它们争夺完全相同的资源,例如食物、水源、领地和繁殖机会。然而,不同物种之间依然存在复杂的竞争关系,只不过形式更加多样化。
以麻雀和家燕为例:二者同为小型鸟类,常常选择重叠的领地筑巢,并且都以昆虫为食。对于有限的筑巢空间和食物资源,麻雀和家燕会彼此排斥,甚至直接在争夺巢穴时发生冲突。这种“干扰型竞争”促使它们在生态位上产生分化,例如在进食时间、筑巢位置等方面寻求微妙的差异,以减少直接对抗。
除了直接争夺外,不同物种之间也会进行“资源利用型竞争”。比如红松鼠和灰松鼠,它们都以坚果为主食。如果两种松鼠生活在同一片森林中,灰松鼠体型更大、耐饿能力更强,于是逐渐取代了红松鼠在许多地区的生存空间。这类竞争通常是间接的,通过更高效地利用资源使对方数量下降。
为什么许多捕食者不会选择捕食同种?例如在狼群中,狼更倾向于群体合作捕猎鹿等大型猎物,而不是攻击自己的同伴。进化上讲,同类之间的攻击风险极大,一旦失手损失的将是宝贵的繁殖者和合作者;进化稳定策略会导致同种个体间建立竞争规则(如争夺地位时的威吓、表演),尽量避免致命冲突,从长远看更有利于种群的整体繁荣。
假如有极端个体偏离这种行为,比如出现一只“专门袭击同类”的狼,它可能短期获利,却因同类报复或失去群体协作而付出更高代价。类似地,在被捕食方中,若某种鸟突然演化出攻击天敌的“反击型”行为,付出能量往往大于获得,最终会因不适应整体策略而被淘汰。因此,无论是种内还是种间,竞争和冲突的“度”都受到进化制约,引导群体逐渐形成最适宜的生态平衡。
基因的合作
传统上说到竞争,我们常常关注动物之间的生存斗争。但事实上,生命体内更深层次的“合作”也同样关键。在每一个生物体内,无数基因必须协同工作、彼此配合,才能支持复杂的生命活动。如果基因各行其是,不仅无法共同达成目标,反而可能引起耗损与冲突,影响个体生存。
我们可以把基因协作比作一个建筑项目的团队。每个基因就像不同的工种,有的负责“打地基”(遗传骨架)、有的负责“布线管道”(调控信号),还有的主攻“装修美化”(表型表达)。如果泥瓦工和木匠都很强,但没有设计图纸或者协调沟通不畅,结果只能是项目失败。同理,某个本身很优秀的基因,如果与整体“基因团队”脱节,反而可能拖累适应度。
下面列举了不同“基因工种”如何合作:
这些基因如果比例、分工搭配不合理,哪怕个别基因很“优秀”,整体的效果也会大打折扣。例如,牛的胃内有分解纤维素的酶相关基因、也有负责营养吸收的基因。如果只有前者没有后者,消化效率不会提升,必须两者配合、比例统筹。
这种基因合作就像建房子招人,不是“越多越好”或“某一种越强越好”,而是各类型、各岗位达到平衡。理想团队可能需要:1位设计师、2位木匠、2位泥瓦工、1位电工,如果全是木匠,房子也是无法建好的。现实中,当某种“岗位”短缺时(如控制新功能的基因),它的适应度价值就会上升,成为“抢手货”;而当这种岗位增多,其他基因类型反而变得稀缺。基因间的互补、协作由此保持,并持续在演化中动态调整。
基因库可以被视为基因的“生态环境”。“好”基因不是静态绝对的,而是指它能在全体现有基因背景下,最大程度提高个体生存与繁殖的概率。
稳定的基因库
长期演化下,不同基因通过反复竞争、尝试和淘汰,最终达成一种“进化稳定”的协作状态。这种稳定状态下,基因间组合彼此适配、配合高效,新出现的变异基因很难单靠自己打乱整个平衡。大部分新基因(不论是突变、重组还是引入)如果难以融入原有基因网络,都会迅速被自然选择淘汰掉,而不会长期存留于种群中。
我们可以把基因库稳定性用团队引入新成员的情况来比喻:
例如,抗病、繁殖、代谢等环节,基因如环环相扣齿轮。即使一个新变异提供了某方面优势,但如果不能与其它“齿轮”啮合,也难发挥整体功效,反而被“刷下来”。正因如此,所谓“优良基因”往往依赖于它所处的网络及组合。
偶尔,也会出现“黑马基因”——它能促进或改造原有组合,最终迅速扩散进整个种群。这类突破性变异往往短时间内打破稳定,导致“基因库大洗牌”。但群体经过适应调整后,会再度进入一个新的、相对稳定的平衡点。这个过程很像“适应度高原”模型:大部分时间平稳、偶尔发生快速跨越。
上图比较了两种主要进化模式:间断平衡模式(蓝线)与渐进演化模式(绿线)。间断平衡理论认为,大部分基因/性状会在长时间内保持不变,偶尔在短暂期内激烈变化——如同团队多年配合突然被新成员“激活”,瞬间蜕变升级。与之相对,“渐进模式”认为性状缓慢、线性地积累改变。
实际上,进化稳定策略(ESS)往往更支持间断平衡:当群体整体处于某种稳定策略时,外来新基因/性状很难单独入侵,系统表现为长期平稳,只有极少数情形下才发生“跳跃”式的突破。
这种“稳定—突变—新平衡”的节律,正是自然界物种多样性演化的真实写照。不仅出现在基因层级,也扩展到物种、生态系统、文化等更高复杂度的系统。
总结
通过进化稳定策略(ESS, Evolutionarily Stable Strategy)的概念,我们能够深入理解为何自然界中的竞争和冲突往往是“适度的”:不是因为动物具备道德观念,更不是出于对“物种利益”的无私关心,而是源于进化过程中——那些选择采用稳定策略的个体更有可能留下后代。
进化稳定策略不仅在动物行为中体现出来,还深入到基因、种群、甚至整个生态系统的结构中。它综合考虑了冲突的代价、对手的策略、环境的动态变化和复杂的互动关系,最终塑造了我们今天在自然界观察到的丰富多样、生动合理的行为模式。可以说,稳定性和协作性是生物世界长期演化所追求和自然筛选出来的最优解之一。