雌雄之间的博弈
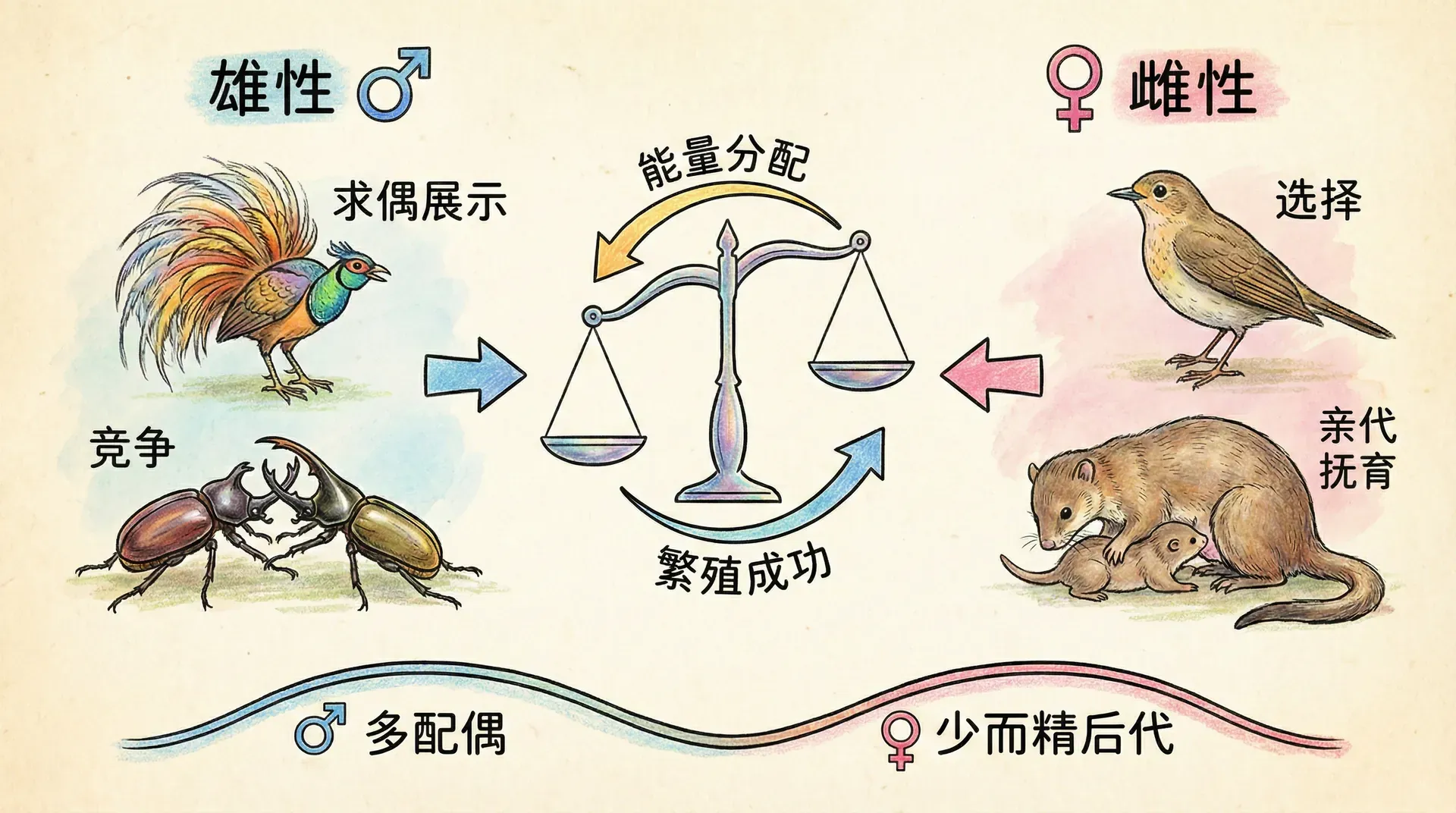
在自然界中,雌性和雄性之间存在着根本性的差异。这种差异并非来自外表特征,而是源于一个更基础的生物学事实——生殖细胞的大小。雌性产生的卵细胞体积庞大、营养丰富,而雄性产生的精子则微小轻便、数量惊人。
一个鸡蛋重约50克,含有丰富的蛋白质、脂肪和各种营养物质,足以支撑一个胚胎发育数周。而一个精子的质量仅有约0.000000005克,比鸡蛋轻100亿倍。这种悬殊的差异决定了雌雄两性在繁殖策略上的根本分歧。
配子大小的差异是理解两性行为差异的关键。雌性每个卵细胞投资巨大,而雄性每个精子成本极低,这导致了截然不同的繁殖策略。
这种差异是如何演化而来的呢?在远古时期,生物的生殖细胞可能都是相同大小的,我们称之为同型配子。任何个体都可以与其他个体融合形成新生命。但随着时间推移,一些个体开始产生稍大的配子,为后代提供更多初始营养,这提高了后代的存活率。然而,这也打开了被“剥削”的大门——另一些个体开始产生更小、更灵活的配子,专门寻找那些大配子融合。小配子的制造者可以生产更多数量的配子,从而获得更多后代。
这种分化一旦开始就无法停止。中等大小的配子处于最不利的位置,既无法提供足够营养,又无法在数量上竞争。最终,两种极端策略胜出:一方产生巨大的、富含营养的卵细胞,另一方产生微小的、高度机动的精子。前者成为雌性,后者成为雄性。
性别比例的进化平衡
既然一个雄性理论上可以让100个雌性受孕,为什么自然界中雌雄比例通常接近1:1呢?从种群角度看,这似乎是一种浪费——那些多余的雄性消耗资源却不参与繁殖。
答案在于个体层面的基因竞争。假设某个种群中雌性过多,那么生育雄性后代的父母将获得巨大优势,因为他们的儿子可以与众多雌性交配,产生大量孙辈。相反,如果雄性过多,生育雌性后代的父母将占优势,因为女儿更容易找到配偶。这种动态平衡最终稳定在1:1的比例。
假设某个地区因为某种原因导致男女比例失衡:
在这个模拟中,当男性比例过高时(比值大于1),生育女儿的家庭在基因传递上更有优势,因为女儿更容易找到配偶并生育后代。这种选择压力逐渐将性别比推回平衡点。
进化稳定的不是数量比例,而是投资比例。父母应该在儿子和女儿身上投入相等的总资源。在大多数情况下,这意味着生育相同数量的儿子和女儿。但在某些物种中,如果养育一个儿子需要三倍的资源(比如海象,雄性需要长得更大才能在竞争中获胜),那么生育三个女儿和一个儿子的策略可能是稳定的。
亲代投资的不对称
由于卵细胞比精子大得多,雌性在受孕那一刻就已经投入了更多资源。这种初始投资的差异产生了深远影响。如果雄性在交配后离开,雌性面临艰难选择:要么独自抚养后代,承担全部成本;要么放弃后代,但这意味着浪费已经投入的昂贵卵细胞。相比之下,雄性的损失要小得多——他只是损失了廉价的精子。
这种不对称导致了一个残酷的现实:在许多物种中,抛弃配偶的往往是雄性。在哺乳动物中尤其明显——雌性不仅提供卵细胞,还要在体内孕育胎儿,分娩后还要哺乳。整个过程中,雄性的生理投入可能仅限于交配那几分钟。
让我们用数据来量化这种投资差异。以中国常见的野生动物为例:
从图中可以看出,雌性在配子生产阶段的投入就是雄性的40倍,而在孕育阶段雄性投入为零。这种巨大的差异解释了为什么雌性更倾向于谨慎选择配偶,而雄性更倾向于广泛交配。
然而,并非所有物种都是这样。在某些鸟类中,雌雄双方共同筑巢、孵蛋、喂养雏鸟,投资相对平衡。在少数鱼类中,雄性甚至承担了更多养育责任。这种差异与物种的生态环境密切相关。
抛弃配偶的时机也存在博弈。有时候,先抛弃对方的一方反而占优势,因为被留下的一方面临更大的沉没成本,不得不独自完成养育。这是一种冷酷但符合基因逻辑的策略。
雌性的反制策略
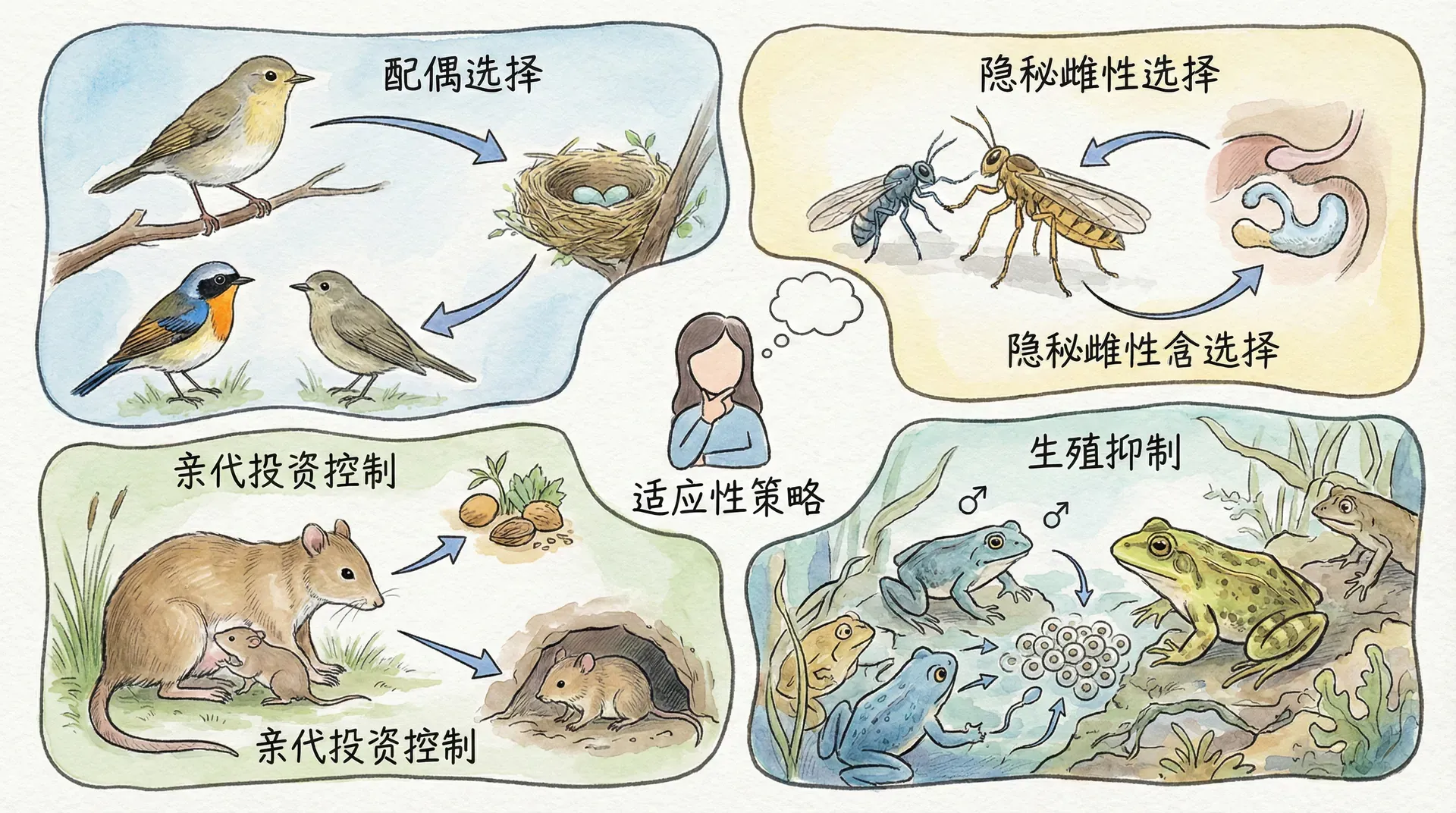
在面对雄性的“剥削”倾向时,雌性其实并非毫无反抗的手段。得益于卵细胞的稀缺与精子的过剩,雌性掌握着重要的“配偶选择权”——简单来说,就是雌性可以对交配提出更多要求,主动筛选、淘汰不可靠的雄性。雌性正处于“卖方市场”,能够通过多种方式提升自身和后代的繁殖利益。
策略一:家庭型伴侣
这种策略的核心是通过考察和筛选,让雌性找到更有育幼责任感的雄性。雌性通常通过以下几个手段提高雄性“逃跑成本”:
- 延长求偶期:借机考察雄性的耐心、忠诚与意愿。
- 实质性前期投入:如要求雄性先建巢、送“聘礼”,或展示劳动能力。
- 共同抚育后代:只有通过上述考验的雄性,才有机会获得交配与参与后代养育。
例如,在中国的朱鹮种群中,雌性会通过长达数周的考察期来筛选雄性,包括筑巢、采集食物等任务。只有表现出持续投入和责任感的雄性才可与之交配。雌性通过这些策略,最大化获得雄性的长期繁殖投资,使后代成长的成功率更高。
燕子和织布鸟的雄性都需要先建造精美的巢穴,雌性进入实地验收后才决定是否与其交配。以下为部分鸟类的前期“家务考察”方式:
值得一提的是,雌性对雄性提出的要求往往更实用:与其“屠龙”、“赛跑”这种炫技,不如筑巢、建房这种真金白银的投入,对后代成长更有直接帮助。
策略二:追求强者基因
若无法期待雄性长期参与养育,雌性可转而追求优质基因或特殊特征,从而确保后代生存优势。这些策略主要表现在:
- 偏好夸张的外貌特征:如孔雀的长尾、狮子的鬃毛、鹿的角等都是性选择形成的显著标志。
- 鼓励激烈的雄性竞争:只有优胜的“强者”能获得大多数交配机会,由此后代基因质量上升。
- “一将功成万骨枯”,牺牲普通雄性的繁殖机会,集中资源扶持极少数优胜者。
以中国南方孔雀养殖场为例,雌性孔雀普遍只青睐尾羽更长、更鲜亮的雄性。尾羽夸张的雄性往往能获得80%以上雌性的青睐,其余雄性则常年“打光棍”。
但问题随之而来:有些特征一开始代表真实能力,比如肌肉结实、动作敏捷。但特征一旦成为雌性的集体偏好,本身就能“自我加强”:即使只是漂亮,拥有这种特征的雄性也会成为下一代的宠儿,子孙世袭。因此,性选择会导致一些特征“越夸张越好”,最终甚至超离实际生存需要。
进化学中有个有趣的“残障原理”理论——只有真正身体素质优异的雄性,才能承受、维系如“巨大尾羽”这样的累赘。这类“炫耀性浪费”反而为雌性提供了可靠的选择信号:既美观又强壮,才是真正的基因优胜者。
雌性通过灵活的策略,不仅能挑选合作型伴侣,还能筛选顶级基因,让自身与后代在演化竞争中获得显著优势。雌性的“筛选权”和策略多样性,是自然界性选择驱动力的重要一环。
两性差异的表现
基于配子大小差异和繁殖策略的不同,我们可以预测并解释许多两性行为差异:
外表装饰
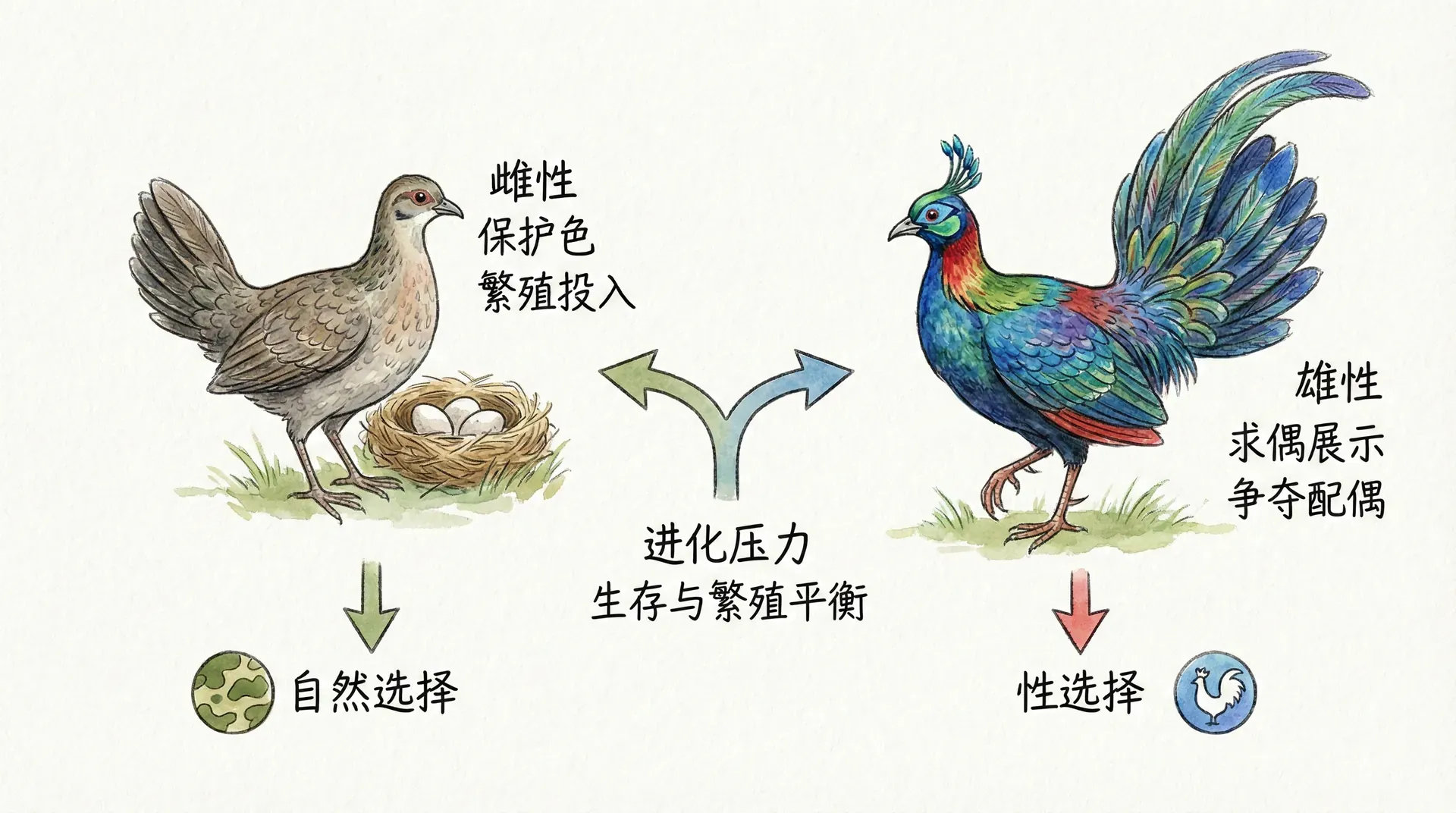
在大多数物种中,雄性比雌性更鲜艳、更具装饰性。这是因为雄性面临更激烈的性选择压力。一个雌性只要产生卵细胞,几乎肯定能找到愿意让她受孕的雄性。但一个雄性如果不够吸引人,可能终生无法交配。
鲜艳的色彩会吸引捕食者,这是一种生存劣势。但对雄性而言,无法繁殖比被捕食更糟糕——前者意味着基因的彻底消亡,后者至少可能在死前留下后代。因此,雄性在“生存”与“繁殖”之间的平衡点偏向后者,愿意承担更大风险来增加交配机会。
在中国的野生鸟类中,这种差异非常明显:
配偶选择的挑剔程度
雌性通常比雄性更挑剔。这不仅体现在同种个体之间,也体现在种间杂交上。一个雌马与雄驴交配产生的骡子是不育的,这对雌马来说是巨大的损失——她投入了11个月的孕期和大量哺乳,最终却没有获得任何基因传递。相比之下,雄性的损失只是一次交配的时间和精力。
因此我们预测,雌性应该更严格地避免种间交配。事实确实如此。在混养环境中,雄性更容易与其他物种交配,而雌性更谨慎。
类似的逻辑也适用于近亲交配。近亲繁殖会增加有害隐性基因纯合的风险,导致后代质量下降。由于雌性在每个后代上投资更多,她们对近亲交配的损失更大,因此应该更严格地遵守避免近亲的规则。如果我们假设年长者更可能主动发起近亲交配,那么父女乱伦应该比母子乱伦更常见,兄妹乱伦应该处于中间水平。这一预测在许多物种中得到了验证。
交配倾向
雄性倾向于更频繁、更随意的交配,而雌性倾向于更谨慎、更有选择性的交配。原因很简单:雄性每天可以产生数百万精子,理论上可以让无数雌性受孕,因此多多益善。雌性产卵速度有限,与更多雄性交配并不能显著增加后代数量,反而可能浪费时间和精力。
这种差异在动物行为研究中反复得到证实。在大多数物种中,雄性表现出更强的交配动机,愿意为交配机会承担更大风险和付出更多努力。
这个图清晰地展示了为什么雄性追求更多交配机会。对雌性而言,一两次交配就足以让所有卵细胞受精,之后的交配几乎没有额外收益。但对雄性而言,每次额外的交配都意味着更多后代,收益曲线持续上升。
人类社会的思考
当我们将这些原理应用于人类时,需要格外谨慎。人类行为受文化影响极大,不能简单归结为生物本能。但某些模式仍然值得注意。
在世界各地的人类社会中,我们确实观察到一些与进化预测一致的趋势:男性平均而言表现出更强的性冲动和更低的配偶选择标准;女性平均而言在择偶时更注重对方的资源和承诺。当然,这些只是统计趋势,个体差异巨大,而且文化因素可以强化或抑制这些倾向。
有趣的是,现代中国社会在性别装饰方面似乎与生物学预测相反:女性使用化妆品、时尚服装等方式装扮自己,而男性相对朴素。这可能反映了人类社会的复杂性——当女性在经济上依赖男性时,她们需要通过外表吸引有资源的男性。但随着女性经济独立性提高,这种模式可能正在改变。
生物学为我们提供了理解两性差异的基础框架,但人类的独特之处在于我们可以通过文化和理性选择来调整这些本能倾向。认识到某些行为模式的进化根源,并不意味着我们必须接受它们,而是让我们更好地理解自己,从而做出更明智的选择。