互助共生
动物世界中,群居行为普遍存在,无论是庞大的鸟群、鱼群,还是草原上的有蹄类动物,乃至社会性昆虫如蚂蚁、蜜蜂,大家都倾向于聚集、协作。这并不仅仅是壮观的自然景象,更是自然选择下的理性结果。群体生活带来了众多优势:比如节省体力、分散被捕食的风险、提升导航与觅食效率。像候鸟选择V字队形飞行,就是通过队形结构利用前方气流来降低能量消耗,鸟群还会轮流更换领飞者,保证个体不会长期过度消耗体力。
相似的协作也出现在鱼群中。许多鱼类通过队形游动,让每只鱼都能“借力打力”,减轻前行阻力,并让捕食者难以锁定目标;而企鹅、野牛等则通过集体聚集抵御寒冷和天敌。在人类社会,合作机制同样处处可见,比如长跑比赛轮流“破风”的兔子、自行车竞速中团队节省体力。这些案例背后的共性在于——只有当协作所得的收益超过单打独斗时,群体行为才能被演化和保留下来。无论对谁而言,协作都不是纯粹的牺牲,而是演化博弈中的务实选择。
群居的核心逻辑是:个体通过集体生活获得的收益必须超过为此付出的成本,这种行为模式才能在演化中保留下来。无论是节省能量、降低捕食风险,还是扩大资源获取空间,只有当合作带来的利大于弊时,群体生活才能演化出稳定的生态模式。
危险区域理论
当捕食者出现时,群居动物会展现出许多令人着迷且自发形成的行为。其中,“危险区域”理论为我们理解群体结构的形成提供了独特视角。
在青海湖畔的藏羚羊群正在草地上觅食,突然一只雪豹悄然靠近。对于捕食者来说,最优策略往往是攻击距离自己最近的猎物,因为这样可以最大程度节省体力,增加狩猎成功率。对于每一只藏羚羊而言,自己周围都会对应一个“危险区域”——只要雪豹步入该区域,这只羚羊就最容易成为攻击目标。
尤其值得注意的是,处于群体边缘的个体,危险区域会比群体中心的个体大得多,因为它们一侧暴露在无人掩护的外部空间。理性的个体为减少自己被猎杀的概率,会本能地向群体内部靠拢,把自己挤进同伴之间,试图将危险区域最小化。反之,中心个体被其他同伴包围,危险来自四周的概率降低很多。正因为如此,动物群体里经常能看到“挤在一起”的情景。正是这些自利的微观动作,促成了宏观上群体结构的紧密和稳定。
即使最初动物分布得很分散,但只要捕食压力存在,每个个体都会主动移动、调整自己的位置,争取钻进同伴之间,把自己从边缘挤进更安全的位置。这种貌似自私的行为,最终却自发促成了群体的形成与集结。没有谁真的在“为群体利益”考虑,但每个体的自利动机和反应,却成就了自然界复杂而壮观的群体行为。这也解释了为何许多动物即使没有高级的协作意识,仍然能够依靠简单的规则演化出紧密协作的群体系统。
警报信号的代价与收益
在四川卧龙自然保护区,野生金丝猴群常常会发出警报叫声。当有成员发现豹子或其他天敌接近时,会发出高频特有的啼叫,提醒同伴迅速隐蔽。这种行为看似无私,因为发出声音后,警报者本身被捕食者察觉的风险显著增加。
事实上,自然界中的警报叫声早已进化出针对性的生存智慧。例如,大多数群居啮齿动物、鸟类和灵长类,都会用刺耳但方向难以判断的高频声发出警报,降低自身暴露的概率。如果请声学专家分析动物的警报叫声,会发现这些声音很难从背景噪音中精确定位来源——这正是亿万年演化压力的产物。
动物为何依然愿意冒险报警?科学家归纳了几类主流解释,下表梳理其核心逻辑与代表案例:
场景举例与分析
1. 亲缘关系驱动
在血缘关系紧密的群体,如金丝猴、狐獴等,个体报警后的风险通过亲属成功逃脱间接补偿自身基因损失,整体有利其遗传利益最大化。
2. “通知策略”:传递群体安全信号
比如在草原上,欧洲田鼠或野兔在发现老鹰时,不是独自藏匿,而是发出警报,整个群体因此同步伏地,显著减少了整体被袭击概率。如果只是自己隐藏,旁边同伴不停活动反倒引火上身。
警报行为并非纯粹利他,而是一种精巧的自保策略。通过让同伴减少动作,发警报者才能真正降低个人暴露几率。
3. “避免脱队”,集体优势
很多社会性鸟类和鱼类更偏向通过警报促进群体整体迁移,如椋鸟和沙丁鱼,一旦有个体发现危险并发出信号,大家会一起起飞或聚集,避免“落单”而成为肉食鸟或鲨鱼的目标。对个体而言,相比单独逃跑,集体逃窜多了额外庇护。
4. 互惠网络与虚假警报
在北美的黑尾草原犬鼠中,警报常常交替出现。科学家发现,警报责任分摊较为均匀时,每个成员都能获益更大。但若有成员长期偷懒不报警,会被群体边缘化。这是一种进化博弈的“合作网络”结构。
动物常见警报方式与伴随风险的演化得失:
草原精灵的跳跃谜题
在宽广的非洲塞伦盖蒂草原与中国内蒙古草原上,羚羊科动物例如黑斑羚(Thomson's gazelle)和黄羊,都以矫健敏捷著称。遇到猎豹或狼群猎杀时,这些动物往往会高高跃起(英文称stotting/pronking),四蹄绷直弹跳至半空,似乎在故意“吸引注意”,挑战直觉的“隐藏才安全”法则。
这种“跳跃”到底有什么进化意义?目前动行为学提出了几种主流假说:
跳跃行为主要解释
-
诚实信号理论(Honest Signal Theory)
动物主动以高难度动作展示体能——“我又快又健壮,你不如去追别的弱者!” 捕食者见状,认为成功率低而选择放弃。大量实验表明,表现出最大跳跃幅度的羚羊个体,被猛兽追捕的概率大幅降低。 -
“扰乱捕食”假说
大幅跳跃使猎捕者短暂迷惑或放弃当前猎物的锁定,有助于个体或集体逃脱。 -
社会信号与配偶择优
犹如运动会上的表演,跳跃也是向同伴展示自身健康与基因实力,增加在族群中的“声望”与异性吸引力。
跳跃高度与获救概率
可以看到,跳得更高的个体,基本不会被捕食;而虚弱或者受伤动物,几乎“难逃厄运”。这说明高难度弹跳的确是个体实力的“硬性名片”。
除此之外,“信号放大器(amplifier)”机制也不可忽略。表面上看,集体高跳行为可能让全群个体都更“显眼”,但实际上,它能帮助体弱者被迅速识别,而强壮者获得安全。整个群体也因此减少了漫无目的的消耗性追逐,节省体力并增加整体生存率。
综上,草原羚羊等高跳“表演”的进化意义十分深远:不仅是个体宣告实力的信号,也优化了群体与捕食者之间的“成本-收益”动态,是动物博弈智慧的精妙体现。
社会性昆虫的合作
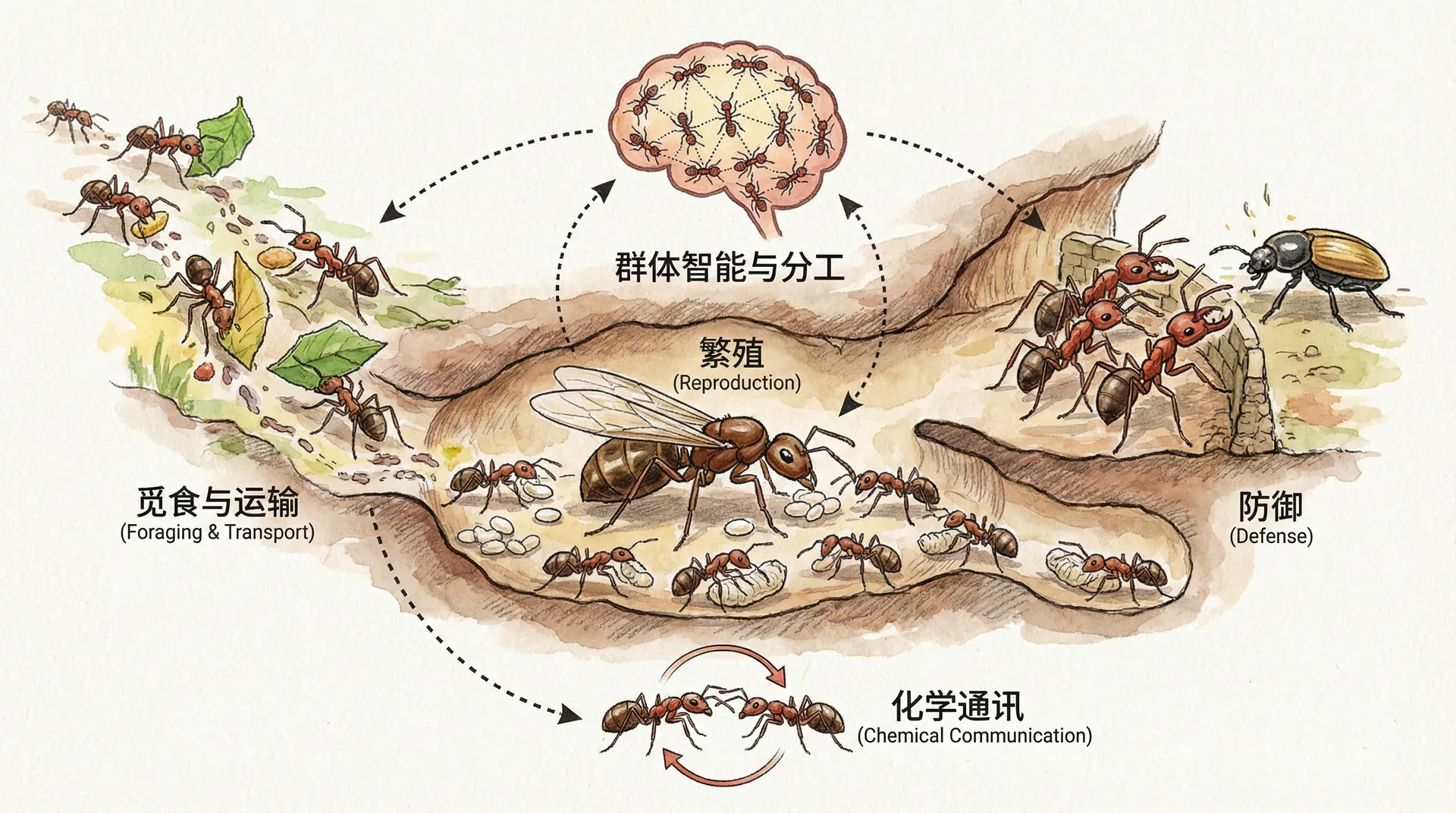
蜜蜂、蚂蚁、黄蜂和白蚁等社会性昆虫,被誉为地球上最成功、最复杂的合作生命体。它们的群落宛如超级有机体,每个成员分工明确、职能高度专一,有些甚至愿为群体牺牲生命。例如:
- 蜜蜂士兵:为保卫蜂巢,可在敌人身上留下毒刺,但自己必死无疑;
- 白蚁战士:进化出巨大脑袋和颚,用于堵住巢穴入口抵抗侵略;
- 蜜罐蚁工蚁:腹部膨大成蜂蜜囊,长期挂在巢顶,成为食物储备库,但失去自主活动能力。
这一切的核心驱动力,是极致的分工。举个分工示意表:
细看工蚁(如蚂蚁或蜜蜂),会发现它们终生忙碌,所做一切皆为群体与蚁后(母亲)服务,几乎没有个体生殖机会。乍看极度“利他”,但从基因层面,工蚁通过间接增加姐妹和母亲的存活繁殖,从而让自己的基因间接获得传承。即“我为群体而活,也为自己的基因存续而活”。
这种极致分工带来的奇迹包括:
- 群体自我组织维持恒定温湿度
- 化学信号高效传递“方向/危险/分工”信息
- 防卫者无惧牺牲“以命相搏”,捕食者难以得逞
更有趣的是,社会性昆虫的“职业”不止于两分。以蚂蚁为例,一巢中往往有工蚁、兵蚁、蜜罐蚁、繁殖蚁后/雄蚁等不同分型,甚至出现专职“奶妈”或“育婴员”,让整个社会运行井然。
社会性昆虫工蚁或工蜂“不育”,其实是进化分工最极致的表现。大规模无私协作,使全体生存概率最大化,是自然界最惊人的社会创新之一。
膜翅目昆虫的遗传密码
蚂蚁、蜜蜂和黄蜂等社会性昆虫,均隶属于膜翅目。它们的性别决定机制与大多数动物完全不同,这是理解其超级合作社会演化奥秘的“遗传密码”。
与普通动物由受精卵发展出雌雄不同,膜翅目昆虫的雄性直接由未受精卵发育得来,身体内只含有母亲的一套基因(单倍体);而雌性由受精卵发育,是常规的二倍体(来自父亲与母亲各一套基因)。此外,雌性发育成工蚁还是蚁后,取决于获得的营养和群体需要而非遗传本身。如此一来,昆虫个体之间的亲缘关系,出现了人类社会难以想象的新奇格局。
具体而言,雄蜂只含有单套染色体,由于没有减数分裂的重组过程,它产生的全部精子遗传信息完全一致。因此,同父同母的姐妹,其父系部分100%相同,母系部分平均有50%基因相同,综合起来,姐妹间的平均基因一致度高达75%,而普通动物无论母女还是姐妹间,通常是50%基因一致度。这意味着,一只工蚁与自身姐妹的相似性,比她与亲生女儿还要高。
从基因自利角度看,帮助“母亲”生产更多姐妹,反倒比自己生孩子更“合算”。这正是为何膜翅目中多次独立演化出了工蚁(工蜂)终生不育、专注养护蚁后后代的现象——她们把自己的生存和劳动全部投放给姐妹,反映了基因“分身”利益最大化的演化妙计。
这种复杂的关系可以用下表理解:
正因如此,膜翅目社会性昆虫成为地球上最成功的社会性动物之一,分工协作和利他行为高度发达。类似的不育社会体系,在膜翅目内部甚至独立起源了十多次,显示出该遗传模式下强大的一致性选择压力。
性别比例的争夺战
由于膜翅目的独特遗传系统,群体内不同“阶层”个体之间对性别比例的期望产生了激烈冲突,这是一场看不见的演化战役。
从蚁后的视角出发,她生产的每一个儿子或女儿,都有50%的基因与自己相同,因此她希望繁殖个体(也就是未来能够繁衍后代的蚁后和雄蚁)的理想性别比例是1:1,资源、能量平均分配。
但工蚁的利益则不同。作为蚁后的女儿,工蚁与姐妹的基因一致度为75%,而与兄弟仅为25%。从基因角度,她们更倾向于协助产生更多姐妹(雌性繁殖蚁),而不是兄弟。这种潜在的基因利益,令工蚁最优策略是在性别投资上采取3:1的偏向——即群体资源的3/4应当投入雌性繁殖蚁,1/4投入雄性。
科学家在不同蚂蚁物种中统计了雌性繁殖个体和雄性繁殖个体的实际投资比例。实验结果显示,被测的20种蚂蚁中,实际资源分配比例与工蚁的“期望值”极为接近,大约为3:1,远高于蚁后的平均1:1比例。这意味着,演化在此博弈中偏向了掌握“婴儿房控制权”的工蚁,她们通过控制幼虫养育流程、摘除或“忽视”雄性幼虫,最终实现了对性别投资的操控。
实际上,工蚁拥有极强的共同行动能力,能够通过分泌特殊信息素,选择性地喂养、照护雌雄幼虫。蚁后试图用遗传和产卵行为调整性别比例的努力,往往被工蚁在实际操作层面“纠偏”。这种复杂的性别分配权争夺,正是社会性昆虫多级利益博弈与演化制衡的精彩范例。
在这种演化机制下,性别比例已不再是“随机”的产物,而成为双方“相互博弈”的结果,展现了基因层面错综复杂的策略较量。
奴役体系的反转
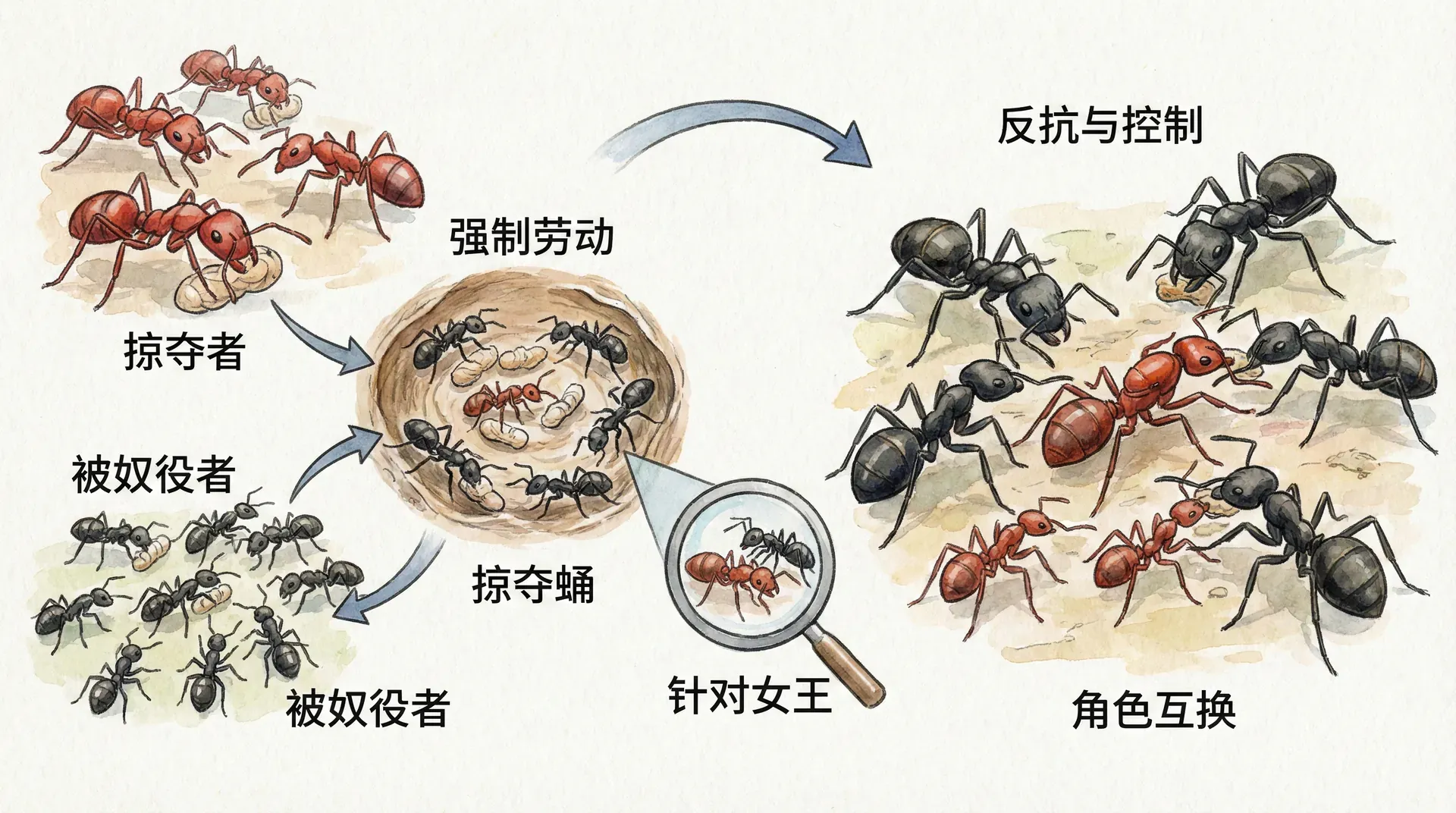
在大自然中,博弈与反博弈从未终结,生物策略的较量常常超乎我们想象。在新疆天山区域,存在一种极为独特的“奴役蚁”体系,其社会结构颠覆了我们对蚂蚁群落的常规理解——这里的“工蚁”不再负责采食、育婴或守卫,而是被进化塑造为专业的“掠夺者”。这些奴役蚁会组队出击,袭击其他蚂蚁巢穴,优先消灭成年守卫,再掠走幼虫和蛹,把“活体劳动力”带回自己的巢中。
这些被掳回的“奴隶工蚁”在新家孵化后,出于遗传本能,会自动融入奴役蚁社会,承担起一切日常维护劳动,包括采集、喂养、清理等。而奴役蚁自己集中精力于掠夺,形成了一种高度寄生化的社会分工模式。
在这一体系下,群体内的性别分配权也出现了反转。因为照料蚁巢幼体的已经变成了没有亲缘基因关系的奴隶工蚁,她们会根据自己物种原有的认知机制,按照默认比例(如3:1高比率照顾雌虫)养育,但这种“偏好”已经指向了奴役者的后代——实际上让蚁后可以绕过原本由本族工蚁掌控的性别控制权。如果奴役蚁蚁后再利用信息素等方式干扰“奴隶”分辨幼虫种类,这种机制就很难被进化修正,因为奴隶工蚁的行为和遗传不再对后代起作用。
长此以往,奴役蚁群体的性别投资将逐步回归传统的1:1分配,蚁后可重新主导后代结构,而被奴役工蚁因缺乏遗传反馈,其“性别照料倾向”无法影响种群后代。这是现实演化博弈中“生产和资源实际操作权”重构的生动范例。
奴役蚁体系演化机制对比
这一现象带来的最大启示,是演化博弈中“实权”常常胜过表面的规则。谁真正主宰幼体培养和资源分配,谁就能决定后代结构——哪怕旧的基因偏好尚未消退,新的生态流程最终也会重塑策略的赢家。权力制衡与策略博弈在自然界轮番上演,只有一时胜负,不见终局。
蚂蚁的农业革命
不仅仅是奴役现象,蚂蚁在合作与创新方面同样令人震撼。例如在我国广东、广西的湿润山区,生活着名为“切叶蚁”的大师级农夫。切叶蚁的地下巢穴可深入3米,拥有上百个房间和迷宫般的通道。令人称奇的是,它们有专门的“蘑菇房”,用来种植特定真菌。
切叶蚁农业系统一览:
- 工蚁采集新鲜叶片,切碎并制成养料基床。
- 在基床上种植专门共生的真菌。
- 真菌分解植物纤维(蚂蚁无法自己消化),蚂蚁收获并食用真菌产物。
- 工蚁定期清理竞争性杂菌,维护真菌“单一作物”。
- 有些蚂蚁甚至会“修剪”真菌,只保留营养丰富的菌体部分。
更神奇的是,非洲的白蚁和新大陆切叶蚁在完全独立的演化路径下,分别进化出了几乎一致的“农业”体系——充分展现了演化中趋同(convergent evolution)的力量。
蚂蚁与蚜虫的契约
田野间还有另一种经典合作实例——蚂蚁与蚜虫的“养殖契约”。在华北、东北平原的田地里,春夏季常见蚂蚁忙碌地照顾植物上的蚜虫。蚜虫能够高效吸食植物汁液,但消化不完全,大量富含糖分的蜜露会由尾部排泄出来。蚂蚁则利用触角刺激,让蚜虫分泌蜜露,自己享用这份“代谢恩赐”。
有趣的是,有些蚜虫逐步演化出更贴合蚂蚁偏好的行为:有的会“憋”蜜露等蚂蚁来“挤奶”;还有些蚜虫尾部形状、分泌物甚至趋向于“模仿”蚂蚁,从而吸引更多的蚂蚁照顾自己。
那蚜虫能获得什么?对比如下:
例如,许多蚂蚁甚至会把蚜虫卵搬回地下巢穴安全越冬,来年春天再搬上植物放养,把“畜牧”进行到底。而被蚂蚁照顾的蚜虫逐步丧失自卫机制,形成彼此高度依赖的一对共生关系。
互惠利他的演化条件
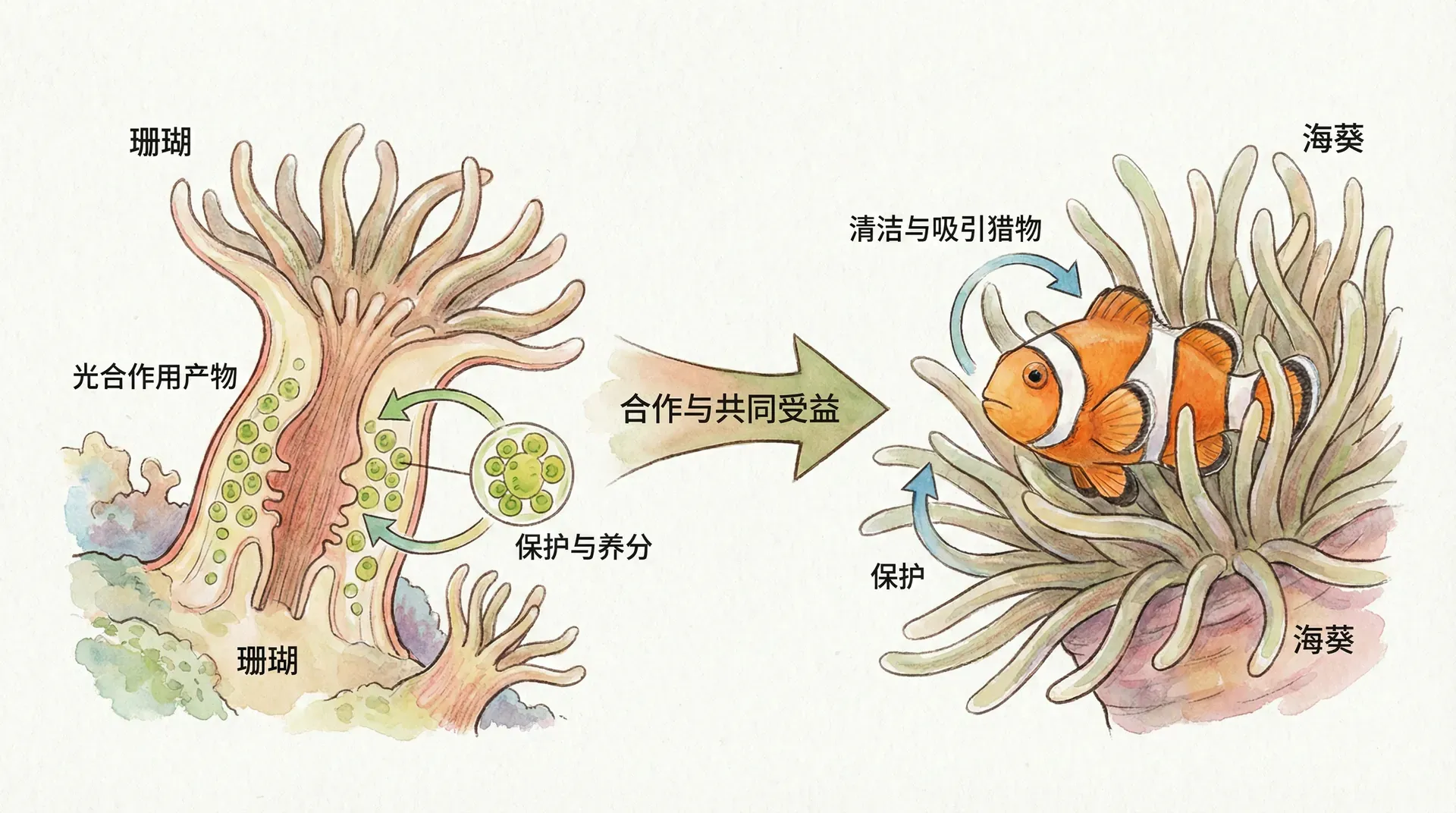
放眼整个自然界,并不是所有共生都能持久,仅在双方能力、资源或需求形成互补时,真正的互利合作才有可能稳定演化。例如蚂蚁和蚜虫:一方善吸植物汁液但无防御力,另一方善于防御却无利用能力。资源交换和专长互补推动了长期合作。
动物与植物、真菌间,也有大量类似互利共生链。比如:
- 地衣:由真菌和藻类高度结合组成。藻类进行光合作用,而真菌提供支持、供水和防护,使双方都受益。
- 珊瑚-虫黄藻:珊瑚为单细胞藻类提供庇护,藻类光合作用产物输送给珊瑚,提高整个珊瑚礁的生产力。
- 蚂蚁与真菌:见前文切叶蚁农业系统。
复杂生命体系本质上就是一张巨大的共生网络:从社区层级(如蚁巢、蜂巢)到每一个细胞内部。比如现代分子生物学已经证明,我们体内的线粒体其实源自被吞噬的远古细菌,成为不可脱离的能量工厂——失去线粒体,任何多细胞动物都无法生存。线粒体至今保留独特DNA和遗传方式,堪称“细胞内共生的化石证据”。
甚至每个基因,都可以视作微型共生单位,依赖其他基因协作运行。整个人体就是庞大基因共生体的集大成。而病毒有可能是这种共生平台中“越界流窜”的基因片段,脱离本有的遗传系统通过空气、体液等媒介传播。把生命看作共生与反共生博弈的超级舞台,有助于我们从全新的视角理解生物演化的本质逻辑。
延迟回报的互惠问题
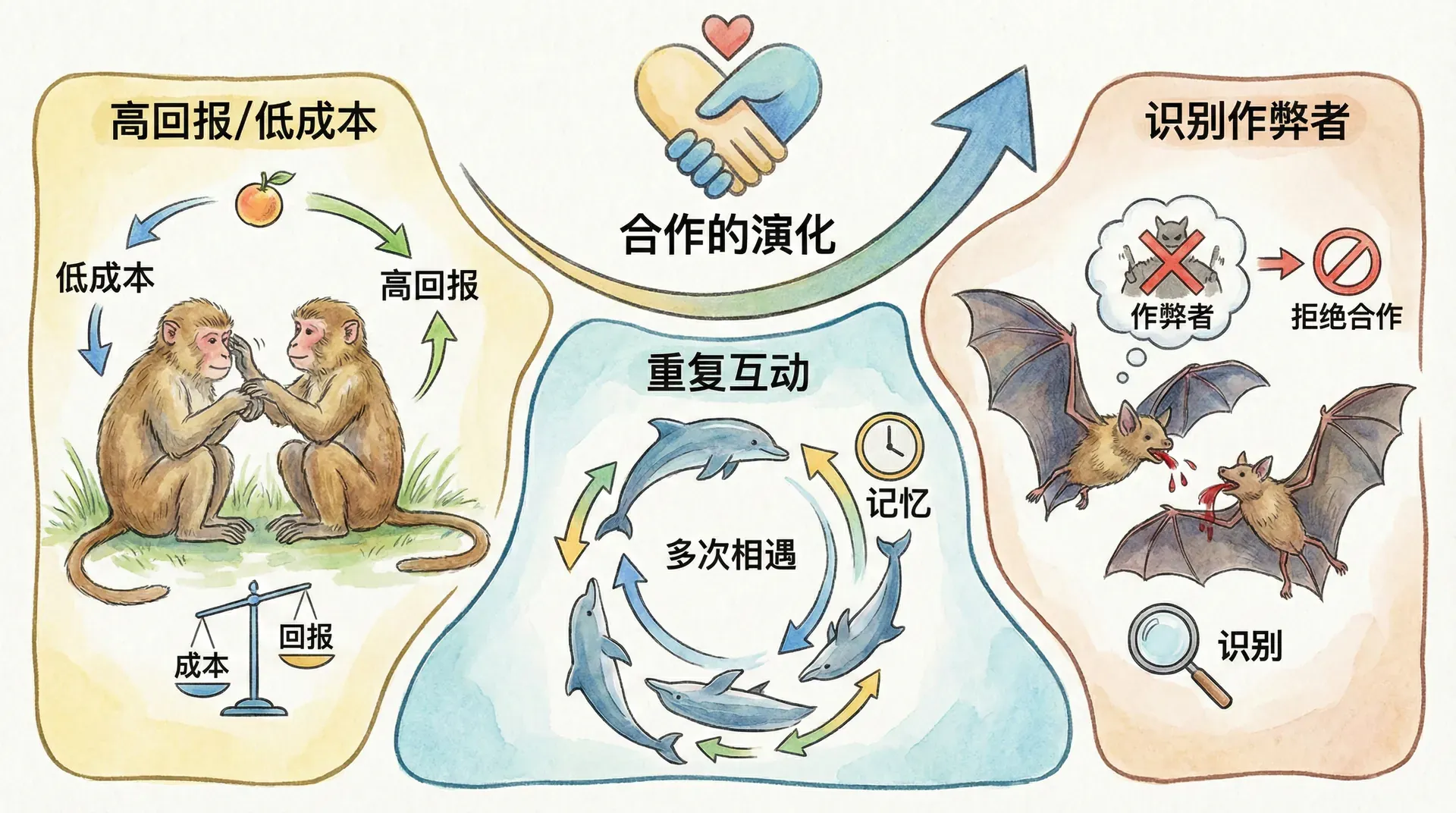
同种个体之间的互惠合作也可能演化出来,只要每一方获得的收益超过付出的成本。当利益交换同时发生时,比如地衣的两个成员持续互惠,演化出这种关系并不困难。但如果施惠和回报之间存在时间延迟,就会出现问题:先获得好处的一方可能拒绝回报。
设想在秦岭山区,有一种鸟类受到携带疾病的寄生虫困扰。每只鸟都能清理自己身上大部分位置的寄生虫,唯独够不到头顶。显然,如果两只鸟互相帮助清理头部,双方都能受益。今天你帮我清理,明天我帮你清理。
这个逻辑看起来很合理,但基因没有预见能力。自私的基因如何演化出这种延迟回报的互惠行为?关键在于:物种必须能够识别和记忆个体。
让我们考虑三种策略在种群中的竞争:
老实人策略:无差别地帮助任何需要帮助的个体。
骗子策略:接受别人的帮助,但从不回报。
记仇者策略:帮助陌生人和曾经帮助过自己的个体,但记住那些欺骗过自己的个体,拒绝再次帮助它们。
在全是老实人的种群中,每个个体都能得到帮助,大家都过得不错。但如果出现一个骗子,它会得到所有人的帮助,却不付出任何代价。骗子的收益比老实人高,骗子的基因会迅速扩散。最终老实人会被淘汰殆尽。
在骗子占多数的种群中,没人帮助别人,每个个体都会因为头部寄生虫而受苦。尽管整个种群面临衰退,但在骗子和老实人的竞争中,骗子依然占优。
但如果记仇者进入种群,情况就不同了。在记仇者和老实人混合的种群中,两者无法区分,都能获得较高的收益。在主要由骗子组成的种群中,单个记仇者处境艰难,因为它要花很长时间才能识别出所有骗子,期间会浪费大量精力帮助不会回报的骗子。
然而,一旦记仇者的比例达到某个临界点,它们之间相遇的概率足够高,能够抵消在骗子身上的损失。此后,记仇者的收益将超过骗子,骗子会被加速淘汰。
在计算机模拟中,我们设定初始条件为:60%的老实人,30%的骗子,10%的记仇者(刚好超过临界比例)。首先发生的是骗子无情地利用老实人,骗子数量激增,在老实人灭绝时达到顶峰。在这个过程中,记仇者也遭受打击,数量缓慢下降。但在老实人消失后,骗子失去了可利用的对象,记仇者开始缓慢增长。随着记仇者比例上升,骗子的生存空间被压缩,最终骗子种群崩溃,记仇者占据统计优势地位。讽刺的是,老实人的存在实际上在早期阶段威胁了记仇者的生存,因为老实人助长了骗子的繁荣。
人类社会中的互惠机制
人类拥有发达的长期记忆和个体识别能力。因此,延迟互惠在人类演化中必然发挥了重要作用。我们的许多心理特征,如嫉妒、内疚、感激、同情等,可能都是被自然选择塑造的,目的是让我们更善于进行互惠,识别欺骗,以及避免被认为是欺骗者。
特别有趣的是那些“微妙的骗子”,他们表面上在互惠,但回报总是略少于所得。人类发达的大脑和数学推理能力,可能正是为了应对这种复杂的社会博弈而演化出来的:进行越来越巧妙的欺骗,以及越来越敏锐的欺骗识别。
货币本身就是延迟互惠的正式化工具。当我们将互惠利他的理论应用到人类社会时,会产生无穷无尽的思考空间。不过,关于这些推论,我们每个人都能够自己思考和判断。