生物进化的历史进程
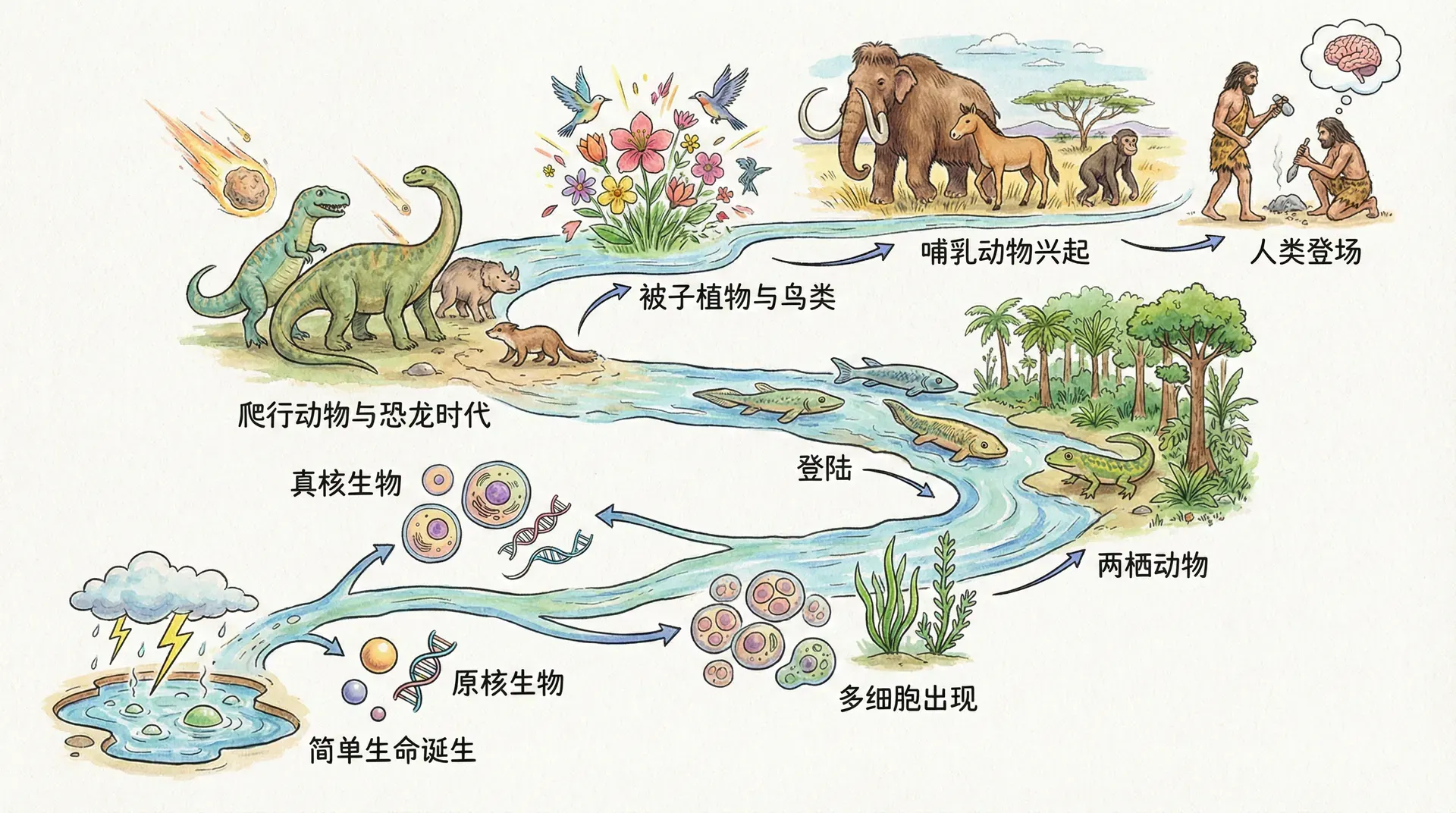
地球生命起源于数十亿年前。当时环境极其原始,最早的生命是简单的单细胞生物,依靠环境里自然形成的有机小分子维持生存。随着可利用有机物的枯竭,仅靠“捡现成”的生物面临危机。这时,一部分生物进化出利用太阳能和无机物(如水、二氧化碳)制造有机分子的光合作用,成为地球上第一批“生产者”;而另一部分则逐步发展为依赖吞噬其他生物获取能量的“捕食者”。这标志着植物与动物两大分支的出现。
此后,光合作用不断优化,推动植物和藻类多样化;而动物方向则专注于摄食和运动能力的提升,演化出丰富的捕食类型。植物和动物这两大类不仅奠定了生命多样性的基础,也推动了地球上能量流动与生态循环的形成。
多细胞生命的出现
随着演化的推进,生物体逐渐从简单的单细胞形式发展为更复杂的多细胞生命。这是地球生命历史上的一次重大飞跃。多细胞生物的出现,使得不同的细胞能够分工合作,专门负责各自不同的功能。例如,在高等动物体内,有的细胞专门负责感知环境(如神经细胞),有的负责运动(如肌肉细胞),有的用于保护(如皮肤细胞),而有的专门承担营养运输、免疫防御等任务。正是这种“分工合作”的机制,使得生物体的结构和功能都变得前所未有的复杂和高效。
每个细胞中都携带着适用于自身分化功能的完整遗传信息副本,但在同一有机体内,细胞会根据不同的“指令”执行特定的基因表达。这使得生物体不仅是遗传信息的集合体,更像是一个高度有序的信息处理与执行系统。细胞之间通过化学信号、物理接触等方式进行沟通,保证整个生物体作为一个整体能精确协调和响应外界变化。
从进化的角度看,个体的统一行为特征是由内部协调机制保障的。尽管遗传信息分布在无数细胞之间,但这种协调能力不是天生完全的,而是在长期激烈的自然选择压力下不断优化的产物。能够维持高效协调的个体获得了生存竞争中的优势,而缺乏内部协调机制的“各自为政”型生物则很难在复杂环境中存续。例如,动物的觅食、逃避捕食者、繁殖等行为都需要全身各个组织和器官的相互配合,任何局部出错都可能导致个体失败。
在日常观察中,我们看到的每一个生物个体都是高度协调的整体。例如一只在树林中奔跑的松鼠,它的四肢、尾巴、眼睛协同工作,这种协调性是数百万年演化的结果。实际上,这种协调不仅体现在外部动作上,还包括内脏器官之间(如心脏、肺、肝脏等)的分工合作,以及体内信号分子的实时调节。
多细胞生命的出现,也为复杂生命活动(如神经系统、免疫系统、器官系统)的演化提供了基础。不同细胞通过“团队协作”,形成了大至动物、植物的各种组织、器官,小到细胞器的精细分工,从根本上改变了地球生态的格局,推动了生命向更高级更复杂的方向发展。
运动能力的革命
动物与植物最显著的差异之一就是运动速度。虽然植物也会运动,比如爬山虎的藤蔓会缠绕攀爬,含羞草会收拢叶片,但这些运动往往是缓慢且不可逆的生长过程。只有在使用高速摄影并加速播放时,我们才能清晰观察到这些变化。而动物的运动速度远远超过植物,可以在瞬间做出复杂且可逆的动作,如跳跃、奔跑、飞行、游泳等。
这种快速而灵活的运动能力,是动物适应竞争激烈的生存环境,寻找食物、逃避天敌、寻找配偶等多种需求的关键。实现这一点的核心装置是肌肉组织。肌肉就像生物体的“动力引擎”,能够高效地将体内储存的化学能(如糖原、脂肪)直接转化为机械能。与蒸汽机或内燃机不同,肌肉产生的机械力是以“拉力”形式表现出来,通过高度复杂的分子结构(主要是肌动蛋白和肌球蛋白的交互作用)将化学能精确转化为收缩运动。这种分子机器的效率和精妙程度,即使是现代工程也难以比拟。
肢体骨骼作为杠杆,肌肉拉动骨骼围绕关节旋转,从而驱动动物完成奔跑、抓握、游泳等复杂动作。此外,不同动物根据生态位和生存策略,演化出了不同类型的肌肉组织:有的专司爆发力,有的擅长持久耐力。
上图展示了人体内两种主要肌肉纤维的收缩特征。快肌纤维能够产生强大的爆发力,适合短跑冲刺;而慢肌纤维则提供持久的力量,适合长时间的耐力活动。短跑运动员通常快肌纤维比例较高,而马拉松选手则慢肌纤维更为发达。这种肌肉结构的差异,也是自然界中动物机能多样性的一种反映。
肌肉运动的复杂性不仅体现在力量和速度上,还包括对空间、时间、力度的精密控制。例如,一只捕食的猎豹需要在高速奔跑时随时调整身体平衡;一只飞鸟在空中作出俯冲、转向等动作时,数百块肌肉与骨骼、神经高度配合,错综复杂却极为高效。这种高超的运动能力,是生存竞争和演化压力共同推动的产物。
神经系统的诞生
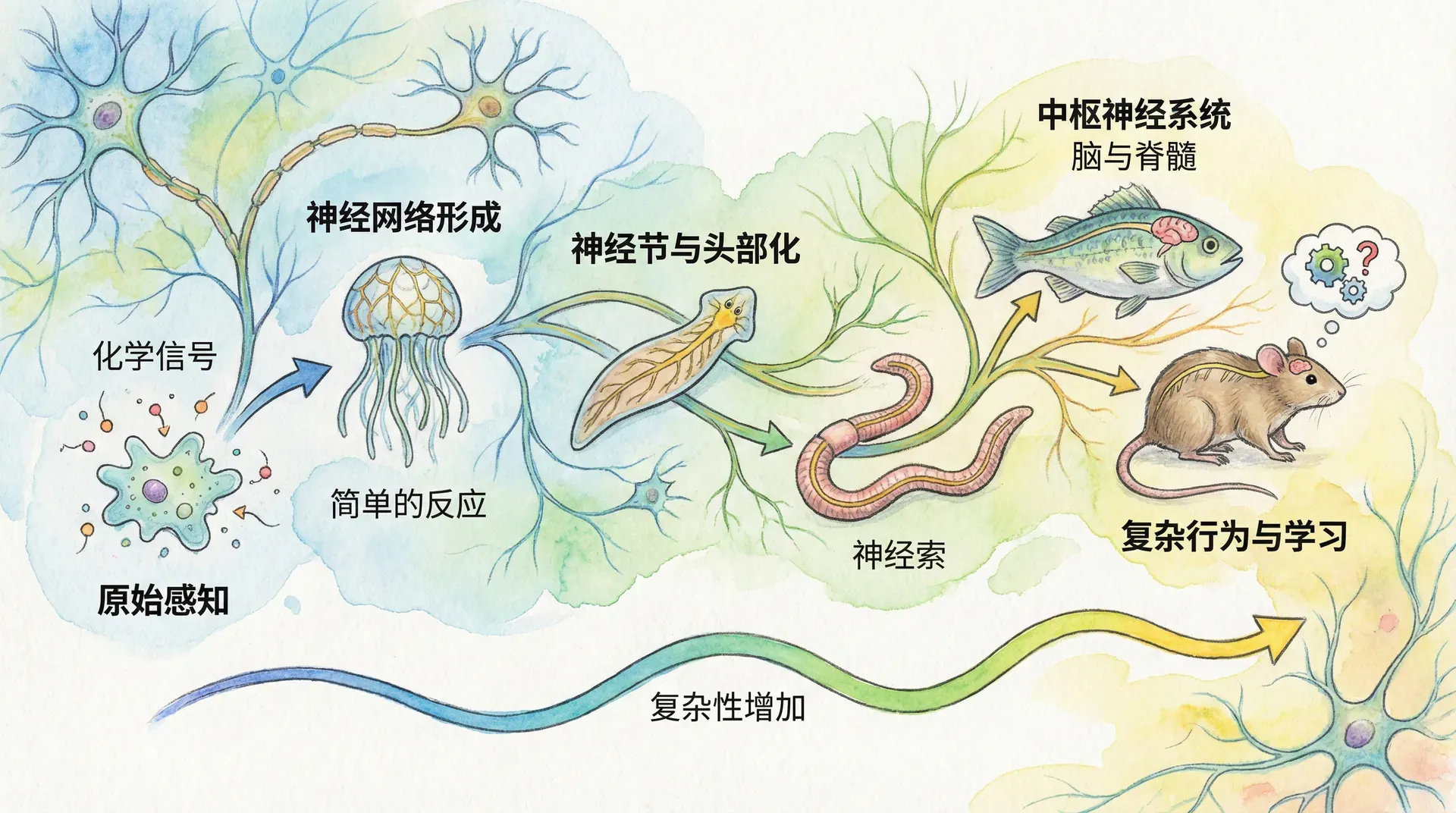
肌肉运动如此复杂,若仅靠简单的物理或化学信号,难以实现精确的时序与空间协调。要高效地控制成百上千块肌肉同步协作,生物体必须发展出更高级的“指挥系统”。于是,神经系统应运而生。这一系统是动物高度进化与复杂行为出现的关键。
在观察现代的人造机械,如自动织布机、工业机器人、生产流水线时,我们会讨论定时与顺序控制的难题。早期机械依赖凸轮或齿轮等硬件结构预设动作顺序,复杂度和灵活性受限。后来的电子计算机利用电流控制开关,实现了高效的信息处理和复杂的序列逻辑。生物体的发展则在数十亿年演化历程中,逐步探索出“生物计算机”—即神经网络。
神经系统的基本单元是神经细胞或神经元。神经元可以把自身收到的信号进行复杂计算处理,再以脉冲的形式输出给其他细胞。这些神经脉冲类似电子计算机中的“数字信号”,能够支持极其复杂的模式识别、信息整合与传递。不同于晶体管那种单一开关,神经元之间通过数万个突触相连,能实现多路信号的并行处理。
人类大脑包含约一千亿个神经元,每个神经元有可能与数千乃至上万个其他神经元通讯。虽然单个神经元的信号传递速度远不及晶体管,但神经网络依靠庞大的并行与冗余系统,实现了高水平的信息处理效率。如果要用目前的集成电路模仿人脑规模和能力,其体积、电力消耗、散热等指标根本不可能适应有机体生存环境。
神经元在动物演化早期即已出现。从最简单的扁形虫或刺胞动物,到复杂的哺乳类和鸟类,神经元和神经网络都以不同形式存在。某些低等生物也许多次“独立”发明过类似结构,这显示了神经系统的巨大进化优势。
神经元的结构特征
神经元本质上是细胞,但形态独特。它不仅拥有普通细胞的细胞核和基因组,还发展出长短不一的突起,从而在形态和功能上大大区别于一般细胞。最典型的是“轴突”——一根可长达数米的细胞突起,通过电化学信号将信息从细胞体传送到遥远的细胞或组织。例如,长颈鹿的颈部有单根轴突直接从脑延伸到躯干深处,负责高效迅速的信号传递。
这些轴突常常和其他神经元的突起编织成束,形成粗大的神经,就像电话主干线,纵横交错遍布全身。除长轴突外,还有密集短突起参与局部网络。这些局域的神经元束在动物体内聚集,成为“神经节”。当神经节进一步体积和功能膨胀时,就演化为“脑”。脑的复杂度远高于单一神经结,是感知、整合与决策行为的重要中心,相当于生物体的“中央处理机”(CPU)。
通过神经系统,动物可以将来自环境的刺激快速整合、处理,并做出多样化、适应性极强的响应。这种“生物计算网络”不仅推动了动物王国的繁荣,也是现实世界中最高级的信息处理系统——人脑和意识——的演化前提。
感知与响应系统
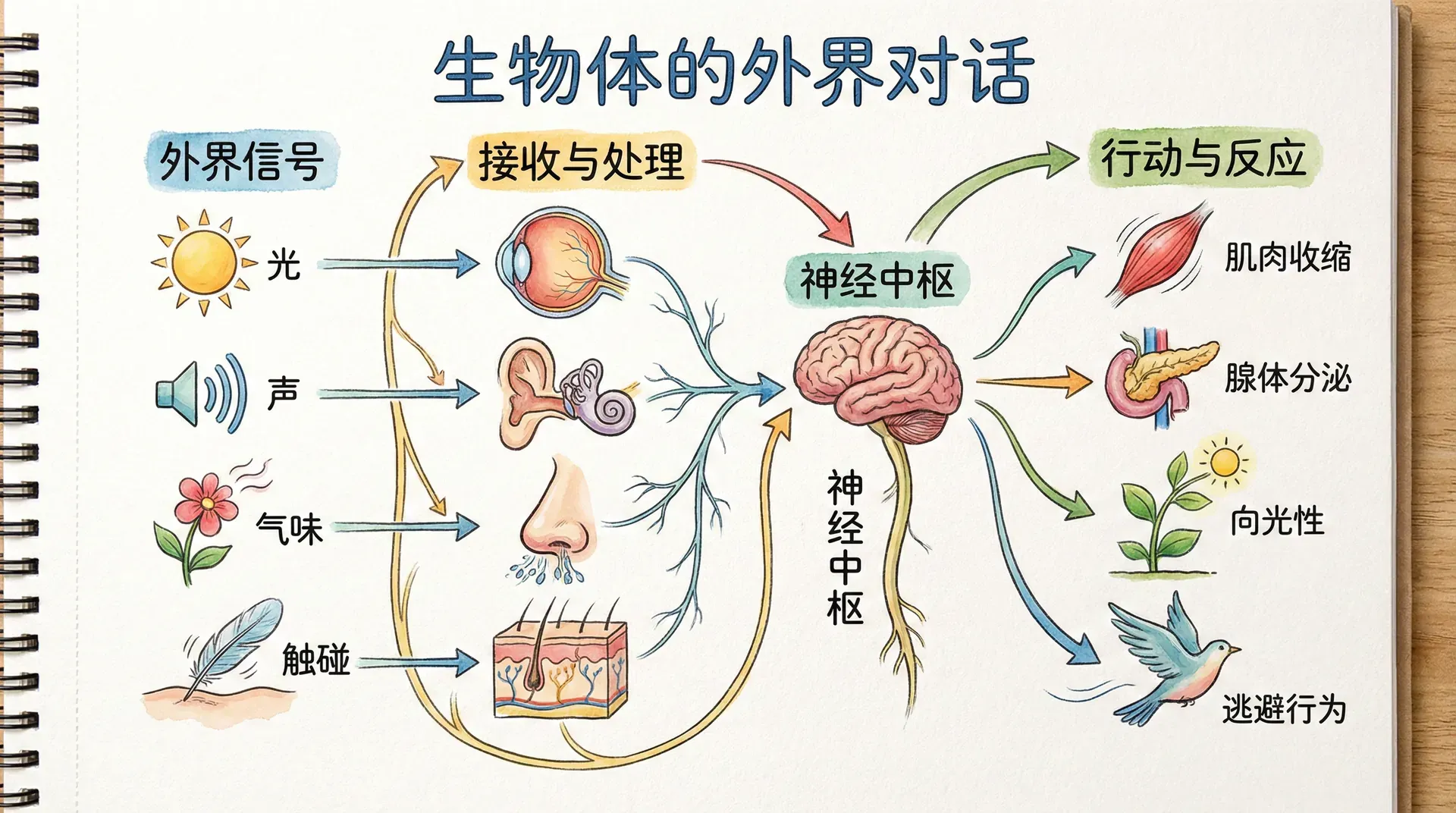
动物能够感知外部世界,并根据这些感知做出精准而高效的反应,这一切都离不开神经系统的精妙协作。大脑通过协调和控制肌肉收缩,促进动物的生存。要实现这一功能,大脑需要与全身肌肉建立直接的传导通路——即运动神经。运动神经如同电缆般传递指令,让手指灵巧弹琴,让猎豹奔跑如风。但行为是否有意义,关键还要看能否及时响应外部的重要事件:只有当有食物时下颌肌肉的动作才有意义,只有面对追逐或逃离目标时奔跑才有价值。也就是说,感知与运动的结合,是复杂动物行为演化的基础。
为此,动物进化出了多样而灵敏的感觉器官,能将外界物理或化学信号转为神经脉冲,供神经系统处理。下表总结了常见感觉器官能感知的信号类型:
大脑通过感觉神经与这些器官相连,实现对环境的时刻监控,使动物能快速做出应对决策。这些感知能力在模式识别与信息整合上远超现有人工智能系统,比如即使戴眼镜换发型,人类也能瞬间认出熟悉的面孔:
在人脸识别中,人类大脑能够在人群中瞬间识别出熟悉的面孔,即使对方改变了发型、戴上了眼镜、只露出半张脸。这种能力经历了数百万年演化,而计算机直至最近才通过深度学习实现类似效果,但依然依赖庞大算力且不及人脑灵活高效。
在生物进化的早期,感觉器官与肌肉反应可能非常直接——如海葵或水螅等低等动物,身体一处受刺激就局部收缩,无需经过中枢神经。但随着活动能力和环境复杂性的提升,更高等动物需要“脑”来整合处理感觉信号和运动反应。这一中介环节确保了来自不同来源的信息能够被综合考量,进而制定更有效、更灵活的行为策略。
记忆的演化意义
随着神经系统层级结构的演化,另一个影响深远的“发明”是记忆。记忆让动物可以将早先的感知、经验和结果存储于神经网络之中,使当下的反应不仅仅依赖当时此刻的感官输入,还能结合过去的经历、教训和关联,做出更加适应性的选择。一个曾经在某块草地中遭遇过危险的动物,后来再经过那片区域时,就会表现出谨慎甚至回避的行动。这正是记忆作用于行为的典型例证。
在数码世界里,计算机的记忆装置(RAM、硬盘)主要用于数据和程序的暂存与调用。虽然计算机的单元记忆极为可靠、稳定,但其灵活性和提取效率,远不及大脑神经网络的复杂结构——后者以大量并行、动态变化的突触连接为基础,能在瞬息万变的环境中进行内容整合、重组与联想。
研究表明,通过反复学习和巩固,信息可以缓慢地从“短期记忆”区转移并固化为“长期记忆”,而不常回忆、巩固的信息则会被遗忘。这就是为什么学习新知识需要多次复习、间隔巩固的原因。在准备考试时,临时抱佛脚虽然有可能让你应付第二天的考试,但很快就会遗忘;而分散在几周时间内有计划的重复学习,则更容易形成稳定而持久的记忆。这一过程对应着神经网络内突触连接的“加固”与“新生”机制,是神经可塑性的基础。
记忆不仅关乎个体学习和生存能力,也是群体行为、文化传承和社会协作的根基。蜜蜂舞蹈、松鸦藏食、灵长类使用工具,都是记忆能力和高级感知-运动整合推动的产物。
目的性行为的本质
在所有动物行为中,最引人注意的一个特征就是明显的“目的性”。我们看到一只动物在“寻找”食物、配偶或走失的小崽时,往往会下意识赋予它主观的“渴望”、“目标”甚至“意图”。我们每个人根据自己的内省经验也知道,在至少某些生物体中,这种目的性发展到了“意识”的层次。
然而,从科学视角理解动物行为时,我们往往避开“意识”这个难以达成共识的哲学议题,转而关注“外在表现足够像有目的的系统”即可。工程学早就实现了不需要意识、但又表现出明显目的导向行为的机器。瓦特蒸汽机的“调速器”正是一个经典例子,它是现代自动控制理论的雏形,也揭示了无意识系统如何展现目的性的本质。
负反馈机制
负反馈机制是工程和生物调控中极其重要的原理。它的基本思想是:系统中有一个测量装置,持续检测当前状态与某个“目标”状态的差距。差距越大,调控装置的行动越强烈,以消除偏差,最终趋于目标。达到目标状态后,调控信号自动减弱甚至消失。这种以“减少偏差”为核心的机制,就是负反馈。生物体在保持内环境稳定(如体温、水盐平衡、血糖)、调节姿势与运动等方面,遍布负反馈回路,其稳定性和适应性远超大多数人造系统。
瓦特调速器是一套与蒸汽机耦合的机械系统。其工作原理:随蒸汽机转速升高,调速器上的两个重球因离心力作用自动被甩向更高位置,带动连杆关闭部分蒸汽阀门,使动力减少,进而令转速下降。当转速降低,重球回落,阀门重新打开,输送更多蒸汽,实现动力恢复。这样一来,无论负载如何变化,锅炉功率输出都能快速回归目标水平,保持机械平稳工作。
上图展示了调速器对蒸汽机运转的巨大影响。没有调速器的蒸汽机因负载变化产生剧烈波动,容易失控损坏。而装有调速器的蒸汽机则能快速且自动地收敛到既定目标值,展现出了类似“有意识自我调节”的效果。这种“目的性”并不是“机器有思想”,而是其内建反馈系统促使其自动优化状态。
生物体运行的众多调节功能——诸如血糖水平、呼吸速率、肌肉张力——也遵循着类似负反馈原理。这些机制帮助动物在不断波动的外部刺激面前,维持重要生理指标的稳定,使其生存概率大大增加。
现代技术进一步“仿生”,实现了各种层次更高目的性的装置。例如制导导弹、自动驾驶汽车、智能机器人,它们可以搜索、追踪、规避、预测目标轨迹,甚至动态修正自身的行动方案,表现得好像有自我意图。尽管这些系统没有半点真实的“意识”,外行人很难不被它们复杂的反应和高度的适应性所迷惑,误以为它们背后必有人的精密遥控。
实际上一切目的性行为,不论是由动物脑还是人工工程完成,内核均是信息感知→误差检测→反馈调节的通用流程。这是理解生物调控和“智能”的基础。
遗传信息的控制方式
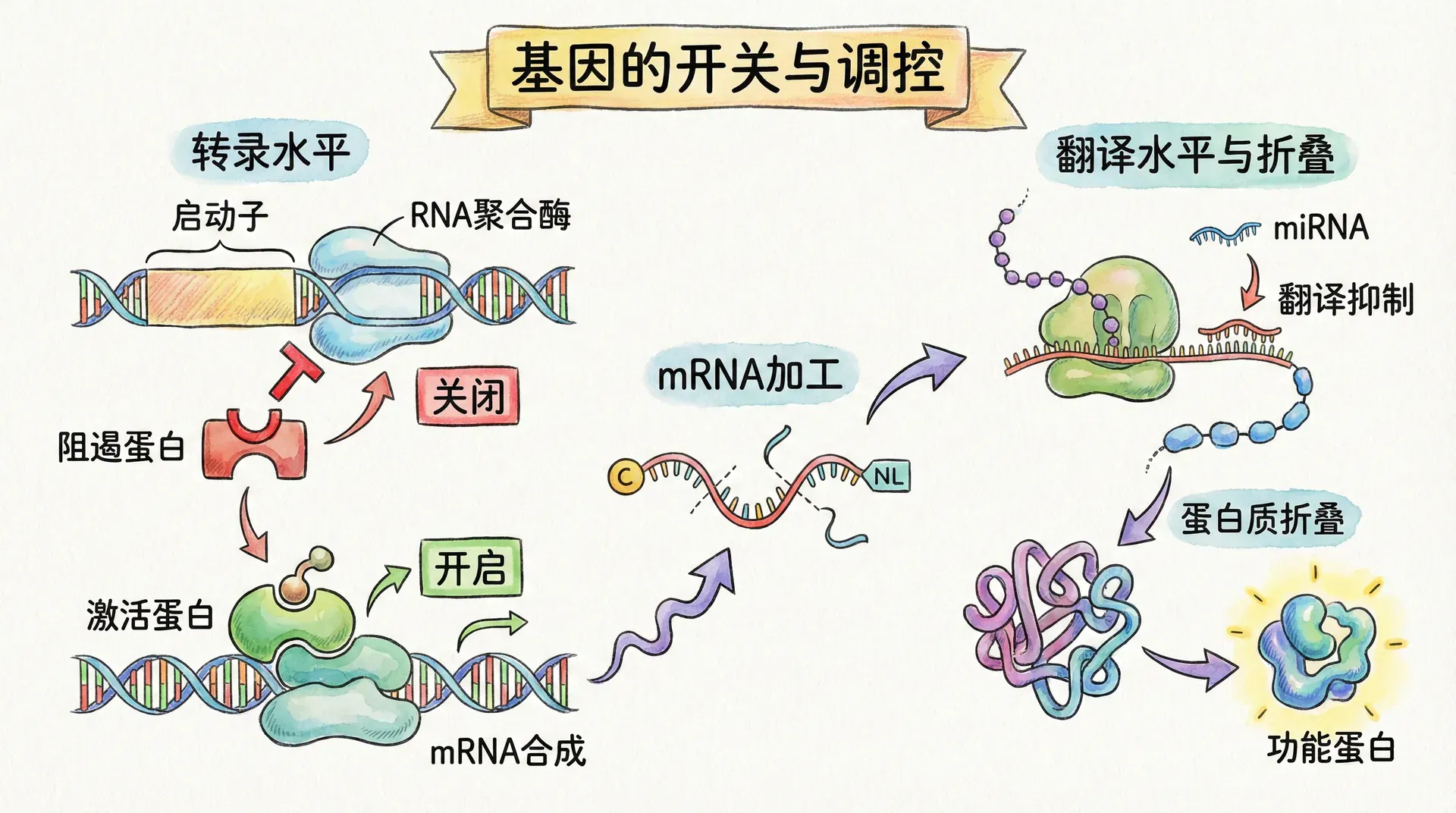
有一个常见的误解:比如因为咖啡机或温控空调等自动化设备最初由人类工程师设计制造,所以它们工作时一定需要人类实时操控。这种观点类似于“智能手机不是会沟通,只是机械执行指令”,其实并不准确。要理解遗传信息的调控本质,就要明白这些观点的问题所在。
现实中,自动化设备如空调、智能家居等,在把参数(温度/时间等)设定好后,人类无需持续干预。工程师或用户设定“目标”,后续调控由内置程序完成,日常运行全靠自动,不必每分钟手动控制。例如你不会总开关空调,设定后系统会自动应对温度变化。
工程师不可能预计家庭中每一刻的温度变化或所有用电场景,也不可能为空调设定针对全部情况的详细剧本。实际中温度变化之多和外部扰动之复杂,难以穷尽。同理,空调自身也无法“提前模拟”所有温度变化再决策。
遗传信息也是以“预先规定规则”而非“时时遥控”的方式影响生物体行为。它更像空调工程师预置的程序:制定好运行原则与反应逻辑,然后让“生存机器”自动去面对环境。遗传信息始终静静存在体内,只有在特定条件下才被激活。为什么不能实时管理?核心原因在于“时滞”问题。
时间滞后的挑战
遗传信息通过调控蛋白质合成而影响个体发育,这是一种从底层改造世界的手段,但过程并不迅速。制造一个胚胎,从单细胞到复杂生物,需要数小时至数月的分工协作。相比之下,行为的本质在于快——捕食、逃生、交流,时时刻刻在毫秒或秒的时标内展开。外部变化即刻触发神经-肌肉的快速反应,基因本身却来不及介入短时事件调节。
假如一家公司总部远在地球一侧,它通过预设指令、培训手册,配置一台自动售货机放置在遥远的他乡。因为物流和通信成本高昂,总部不可能对每次商品出货做出指令,而是将可能遇到的状况、基本操作逻辑和应对优先级一并写入机器。自动售货机面对顾客时,不需要总部联系,也不会每卖一瓶饮料就请示上级,而是按照HQ预先安排的程序自动判定、执行。
现实中,相隔甚远的机器和总部,无法以对话的形式高效操作,只能靠提前预设详细方案。当客户光顾自动售货机、机器遭遇掉电或货品短缺等情况时,只能靠内建程序来应对,而非临时请示。宇航任务中的远程探测器就是如此:探测车登陆火星后,由于几十分钟的信息传输滞后,只能运行具有高度自治的本地程序,并根据之前编写好的规则做出决策,地球总部仅能偶尔下达大方向调整。
同样,遗传信息本身不是一个实时“遥控者”,而是一个“设计说明书”或“自动控制策略”:通过提前设定好逻辑与反应的原则(如:遇见威胁时趋避、饥饿时寻找食物),让生物体内部的“自动程序”负责日常的细节运转。这种“预先安排—自主运行”的机制,才是基因如何“控制”生物机器的真正方式。
遗传信息无法一一预测未来,但能通过内建的程序让生物试图应对最多变的环境。比如探测车会根据温度自动加热、自动分析土壤,无需地球工程师一直遥控。我们的基因同样只能以高度概括性的规则预设各种应激反应,不能事无巨细,而必须用策略性的方式提前赋能生存机器,在千变万化的世界中自主应对。
总结
遗传信息的作用类似“基于过去进行预测”,所有基因实际上都记录了祖先在漫长演化过程中积累的生存经验和对某些环境的适应之道。这些信息通过影响个体的发育、结构和本能行为表达出来。例如,北极熊之所以拥有厚厚的毛皮和白色保护色,就是因为它们的祖先长期处于严寒、冰雪覆盖的环境中,只有具备这些特征的个体才能更好地生存并繁衍后代,因此这种适应性逐渐被固定在基因中。基因本质上是历史的记录者,只能反映过去环境的主导趋势——如果外部世界突然发生剧变,比如气温骤升或者生态系统被破坏,这些原本赋予优势的遗传特征可能反而成为负担,失去原有的保护作用。
然而,复杂多变的环境往往充满不可预见的挑战,仅靠遗传信息中的“过去经验”很难覆盖所有突发状况。为此,许多生物体还进化出了高效的学习机制,能够根据个体自身的生活经历,在生命过程中灵活调整行为策略和适应方式。遗传提供了适应的“基础蓝图”,而学习则像是对蓝图的实时修正和补充,根据现实具体问题随时优化自身反应。遗传与学习的协同作用,使生物能够既承继祖辈适应力,又拥有自我调整和创新的能力,从而在变化莫测的世界中长期持续地生存和发展。