染色体与遗传的多样性
当我们想要深入了解遗传的规律时,很快就会遇到一个实际问题:人类生育周期长,每次只生一个孩子,而且人体细胞中有46条染色体,观察起来相当复杂。直到1957年,科学家们才最终确认人类细胞中染色体的确切数量,在此之前大家一直以为是48条。这说明即便是最基本的观察工作,在人类身上也并不容易完成。
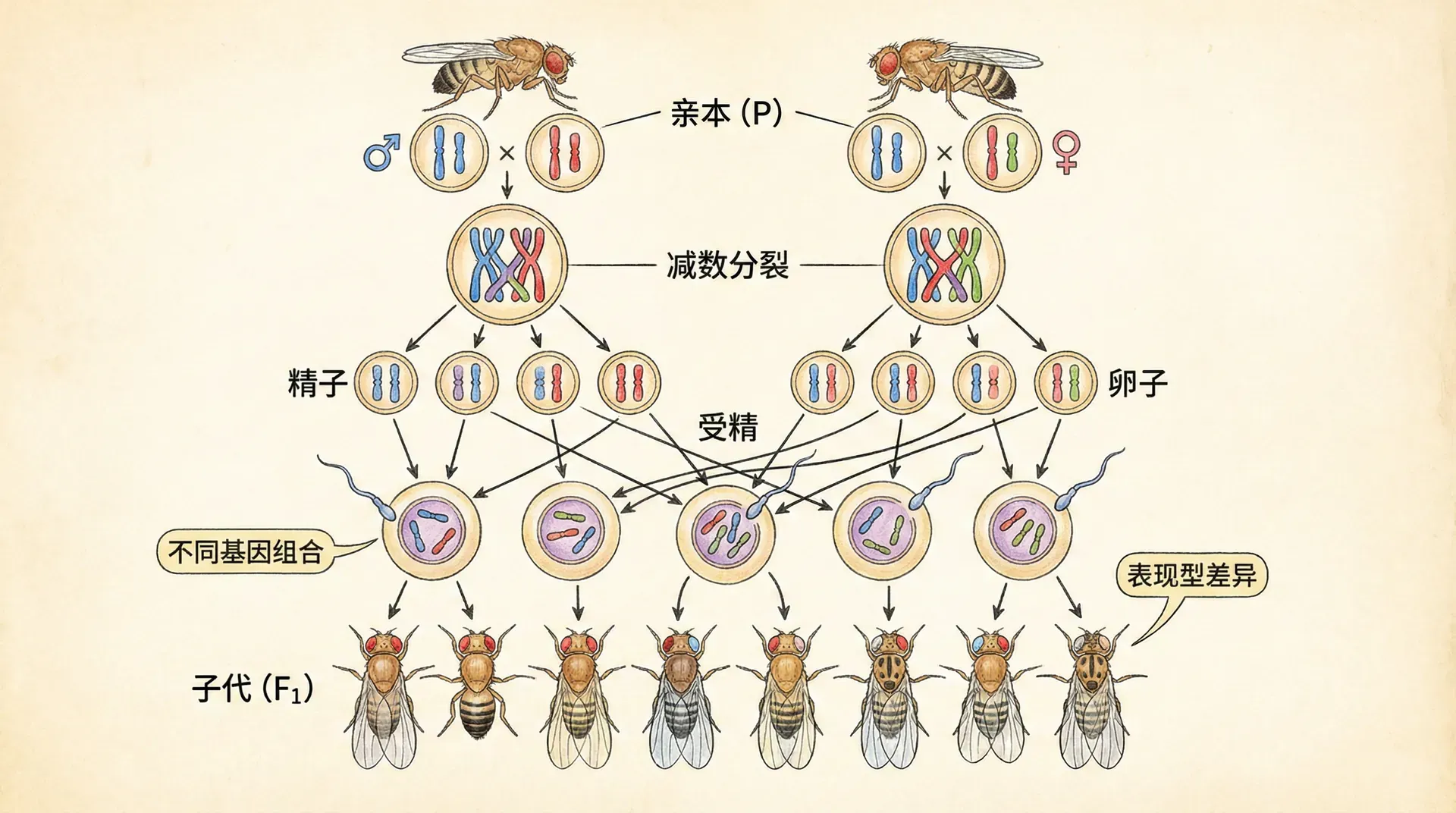
正因如此,遗传学研究需要选择更合适的实验材料。1906年,美国科学家托马斯·亨特·摩尔根开始使用一种小小的昆虫——果蝇进行遗传学研究。这种选择堪称完美:果蝇体型小巧,只有约1毫米长,饲养简单,只需要一个玻璃瓶和一些食物就够了;它们繁殖速度快,大约两周就能产下一批新的后代;最重要的是,果蝇的细胞中只有8条染色体,配子(生殖细胞)中则有4条,这让观察和分析工作变得容易得多。
在随后的半个世纪里,果蝇成为遗传学研究中使用最广泛的模式生物。摩尔根因为在果蝇研究中的杰出贡献,于1933年获得了诺贝尔生理学或医学奖。值得一提的是,中国科学家在遗传学领域也做出了重要贡献,比如谈家桢先生就是中国现代遗传学的奠基人之一,他也曾深入研究果蝇的遗传现象。
染色体的配对规律
仔细观察果蝇的染色体,我们会发现一个十分有趣且重要的现象:这些染色体总是成对出现,并且每对染色体在形态和大小上高度相似。以果蝇为例,它的体细胞中共有8条染色体,表现为2条点状染色体,2条短直线染色体,以及4条较长且中间弯曲呈V形的染色体。这8条染色体可以分成4对 homologous(同源)染色体,每对来自果蝇的父母各一条,彼此对应、携带着相同类型但可能略有差异的遗传信息。
当果蝇进行生殖时,无论是产生卵细胞还是精子,即配子形成时,每个配子只会获得每对染色体中的一条。也就是说,每个配子中都只含有4条染色体,分别为:1个点状染色体、1条短直线染色体,以及2条V形染色体。科学家们从未观察到像“2个点状染色体”或“3个V形染色体”这样的组合出现在单个配子中,这再次表明染色体的分配存在着严格的规律。这种规律性分配被称为“配对分离规律”(Law of Segregation),是遗传学中的基础法则之一。
不光是果蝇,这种染色体成对配对、在减数分裂时分离的规律,其实在绝大多数真核生物中都普遍存在。尽管不同生物的染色体数量、形态差异很大,但“同源染色体成对存在,减数分裂时各自分开进入不同配子”的规律却几乎没有例外。下面的表格展示了几种常见生物的染色体状况:
以上不仅展现了不同物种在染色体数量上的巨大差异,还传递出一个耐人寻味的信息——染色体数量的多少与生物的复杂程度之间,并不存在简单的正相关关系。例如,小麦的染色体数目(42条)甚至超过了人类(46条),但这主要是由于小麦在进化过程中经历了多次杂交和染色体加倍,形成了多倍体结构,而人类则始终维持着二倍体。换句话说,染色体数量并不能用来简单衡量一种生物的“高等”或“复杂”,而更多反映了该物种的进化历史和遗传策略。
实际上,正是这种成对出现、在有性生殖过程中进行严格分配的规律,为后代的遗传多样性奠定了基础。同源染色体在减数分裂时的自由组合,以及随后可能发生的交换(基因重组),共同决定了每一个配子的遗传组成都是独一无二的,为自然选择和进化提供了丰富的素材。这一现象不仅体现在实验室的观察中,也普遍存在于自然界的亿万生物中,是生命遗传多样性的重要源泉。
性别是如何决定的
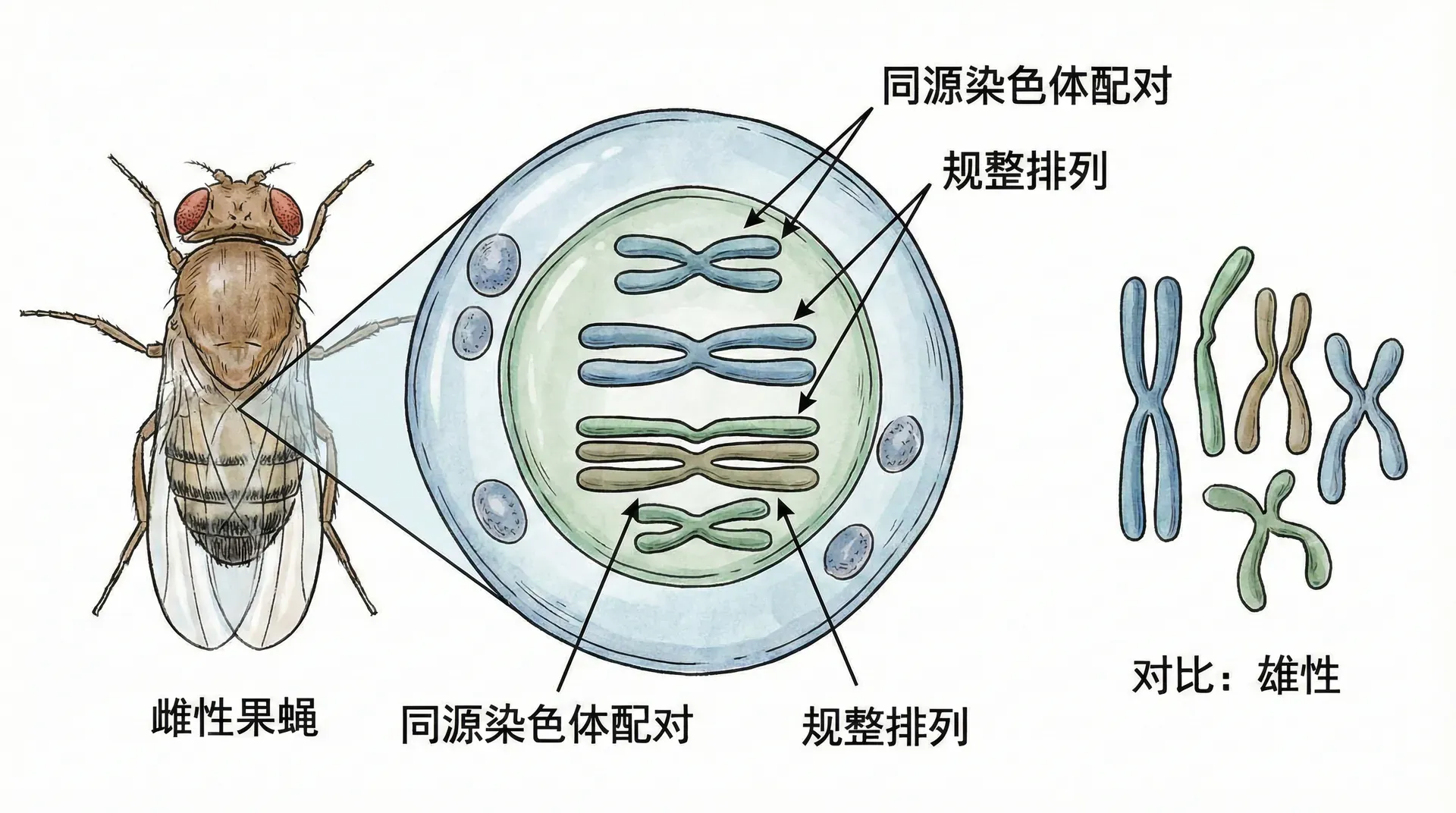
染色体配对现象在雌性果蝇身上表现得最为规整:所有染色体都能找到完全相同的配对。但在雄性果蝇中,情况略有不同。雄性果蝇同样有2个点状染色体和4个V形染色体,但剩下的那对染色体却不完全匹配:一条是直线形,另一条稍长且弯成J形。
1905年,美国动物学家埃德蒙·威尔逊在研究蜜蜂时首次注意到这种特殊的染色体对,并将它们命名为X染色体和Y染色体。这个名字一直沿用至今。雌性果蝇拥有两条直线形的X染色体,而雄性果蝇拥有一条X染色体和一条J形的Y染色体。
这种染色体的差异直接决定了后代的性别。当雌性果蝇形成卵细胞时,每个卵细胞都获得一条X染色体。而雄性果蝇形成精子时,由于存在不匹配的染色体对,会产生两种精子:一半携带X染色体(我们称之为X型精子),另一半携带Y染色体(Y型精子)。
受精时,如果是X型精子与卵细胞结合,受精卵就拥有两条X染色体,发育成雌性个体;如果是Y型精子与卵细胞结合,受精卵就拥有一条X染色体和一条Y染色体,发育成雄性个体。由于两种精子的数量大致相等,受精时哪种精子能成功完全是随机的,因此从长期来看,雌雄后代的比例接近1:1。
人类的性别决定机制与果蝇基本相同。人类的Y染色体相比X染色体要小一些,看起来像个小斑点,但决定性别的原理完全一致。女性拥有两条X染色体,男性拥有一条X染色体和一条Y染色体。根据中国第七次人口普查数据,我国总人口中男性占51.24%,女性占48.76%,性别比例基本符合随机受精的理论预期。
不同物种的性别决定方式略有差异。有些物种中,雄性没有Y染色体,X染色体处于单独存在的状态;而在鸟类等一些物种中,反而是雌性拥有不匹配的染色体对。
基因连锁现象的发现
染色体理论很好地解释了遗传的许多规律和性别的决定机制,但随着科学家对遗传现象的深入研究,发现光靠染色体的“数量”并不能解释所有的遗传现象。原因很直观:细胞内的基因数量远超染色体数量。例如,人类有46条染色体,但却拥有两万多个基因,而可遗传的性状数以千计,比如身高、血型、眼睛颜色、嗅觉敏感性等等。
这里我们不妨用一个比喻:如果把一条染色体比作一串项链,那么大量的基因就像项链上的珍珠。每条染色体可能串着几百乃至上千个“珠子”(基因)。这些基因并不会自由漂移到其他染色体上,往往“拴在一起”,在遗传时作为一个整体被“打包”传递——这就是所谓的基因连锁(gene linkage)。即位于同一条染色体上的基因会作为一个整体一起遗传。
基因连锁现象和特点:
孟德尔提出的“独立分配律”其实是建立在他所选的豌豆7个性状分别在不同染色体上的巧合基础之上。如果当年他选择了珠形和皱形豆荚这样的性状(这两个基因恰好邻近地位于同一条染色体),很可能会发现这些性状总是一起出现或一起消失。这说明“独立遗传”其实只适用于不同染色体上的性状。
我们看玉米中的一个例子:
- 玉米籽粒的颜色(黄色Y显性/白色y隐性)与质地(圆滑S显性/皱缩s隐性)两个基因,若这两个基因紧密连锁在一条染色体上,则在杂交和自交后,后代表现型中主流组合为“黄色圆粒”与“白色皱粒”,很少出现“黄色皱粒”或“白色圆粒”这样的新搭配(除非产生了基因重组)。
类比一下:就像买项链,不能只要其中几颗珠子一样,染色体上的基因常常是“打包出售”的。
交叉互换打破连锁
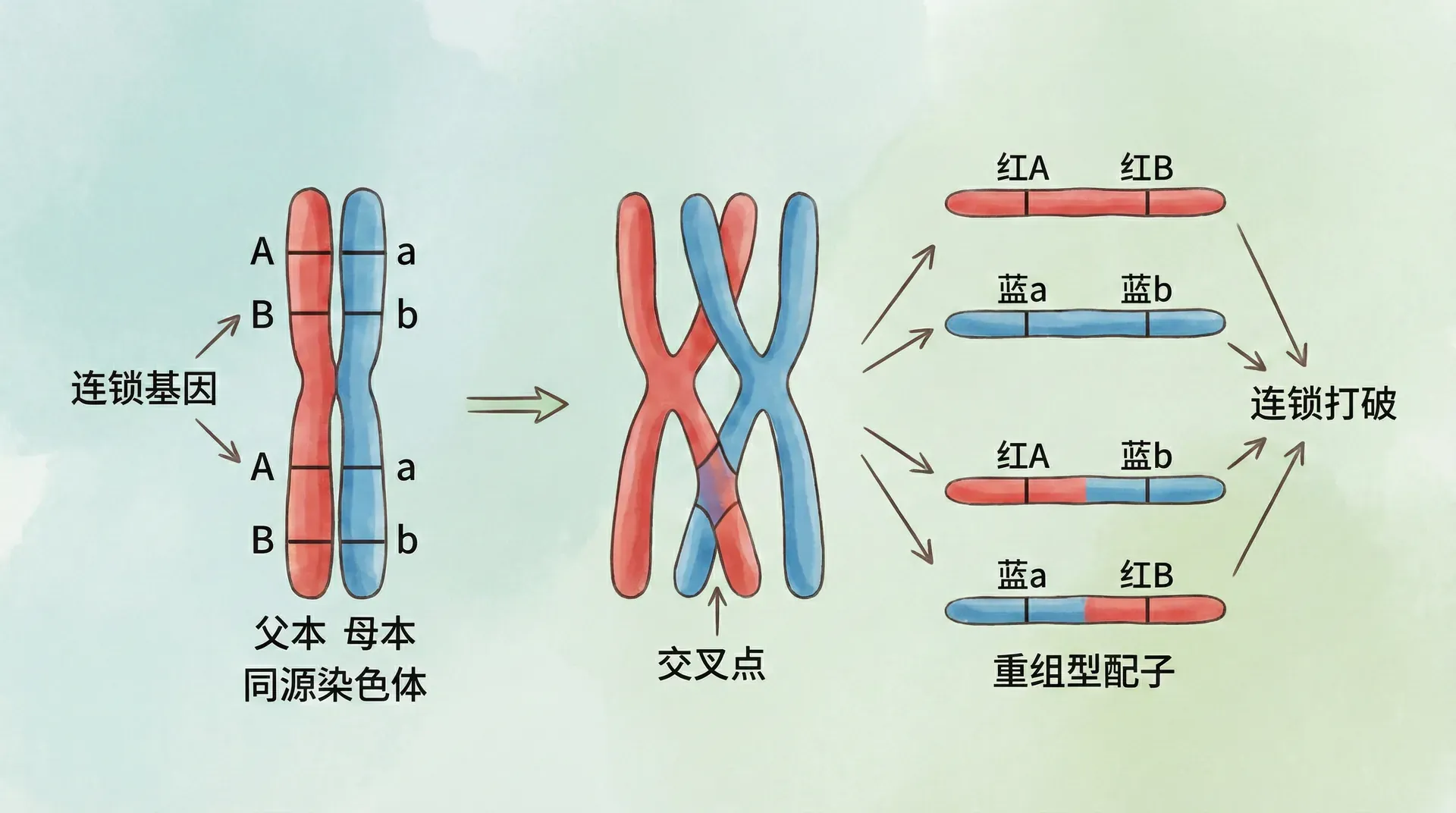
然而,现实中的遗传规律比“全部打包”更灵活。当科学家做杂交实验时,发现偶尔会有后代出现原本紧密连锁基因并未绑定在一起的表现型。例如上面玉米的例子里,还是会少量出现“黄色皱粒”或“白色圆粒”,这说明染色体上“打包”的基因有时会“拆分重组”。
其原因在于——交叉互换(crossing-over)。在减数分裂形成配子的过程中,同源染色体会靠近并有可能发生部分片段的互换。这就好像两条项链相遇时,交换了一些珠子。
下方演示交叉互换对于基因组合结果的影响(以果蝇实验为例):
交叉互换发生后,打破了基因的连锁关系,使得原本连在一起的性状有了新的组合方式。这种现象为基因的多样化提供了额外的可能,也为生物界丰富的遗传变异创造了实际基础。
1911年,摩尔根率先提出可以通过测量不同性状同时分离出现的频率,反推这些基因在染色体上的距离。他打了个通俗的比方:如果两颗珠子距离很远(即两个基因在染色体上距离远),项链从之间断开的概率就大;如果两颗珠子很近,发生“拆分”的概率就小。科学家们正是利用这一点,在1917年绘制出了世界上第一张基因连锁图谱。
这种研究方式理论上适用于所有生物,但在染色体多、繁殖周期长的生物(如人)中,操作较复杂。而果蝇、玉米因其染色体数目不多、种群大和繁殖快,成为经典实验材料。
交叉互换不仅为染色体上的基因提供了更多重新组合的可能,还极大提高了后代的遗传多样性,是自然选择和进化的重要推动力之一。
组合数量的惊人增长
现在让我们算一笔账,看看染色体重组能带来多少种可能性。以人类为例,我们有46条染色体,一个婴儿从父亲那里继承23条,从母亲那里继承23条。
对于第一对染色体,假设父亲的两条分别是A和A',母亲的两条是a和a',那么婴儿可能得到的组合有4种:Aa、Aa'、A'a、A'a'。第二对染色体也是如此,第三对、第四对……一直到第23对,每一对都有4种可能的组合。
那么总共有多少种组合呢?答案是4×4×4×……×4(23个4相乘),也就是4²³。这个数字大约是100万亿(100,000,000,000,000)!
这个图表清楚地展示了组合数量的指数增长趋势。当染色体对数较少时(比如果蝇只有4对),组合数量还比较有限;但随着染色体对数增加,可能的组合数量迅速爆炸式增长。
即使父母的某些染色体完全相同(比如A和A'的基因组成一模一样),也不会明显减少组合的多样性。因为不同个体的染色体几乎不可能完全相同。这意味着,除了同卵双胞胎,任何两个人拥有完全相同基因组合的可能性几乎为零。
更何况,我们还没有考虑交叉互换的影响。交叉互换会让染色体本身也在世代间发生变化,产生全新的基因组合。如果把每个基因的可能组合都考虑进来,人类基因的组合数量将大到难以想象。可以说,每个人都是独一无二的,这在遗传学上有着坚实的数学基础。
多倍体与基因的协同作用
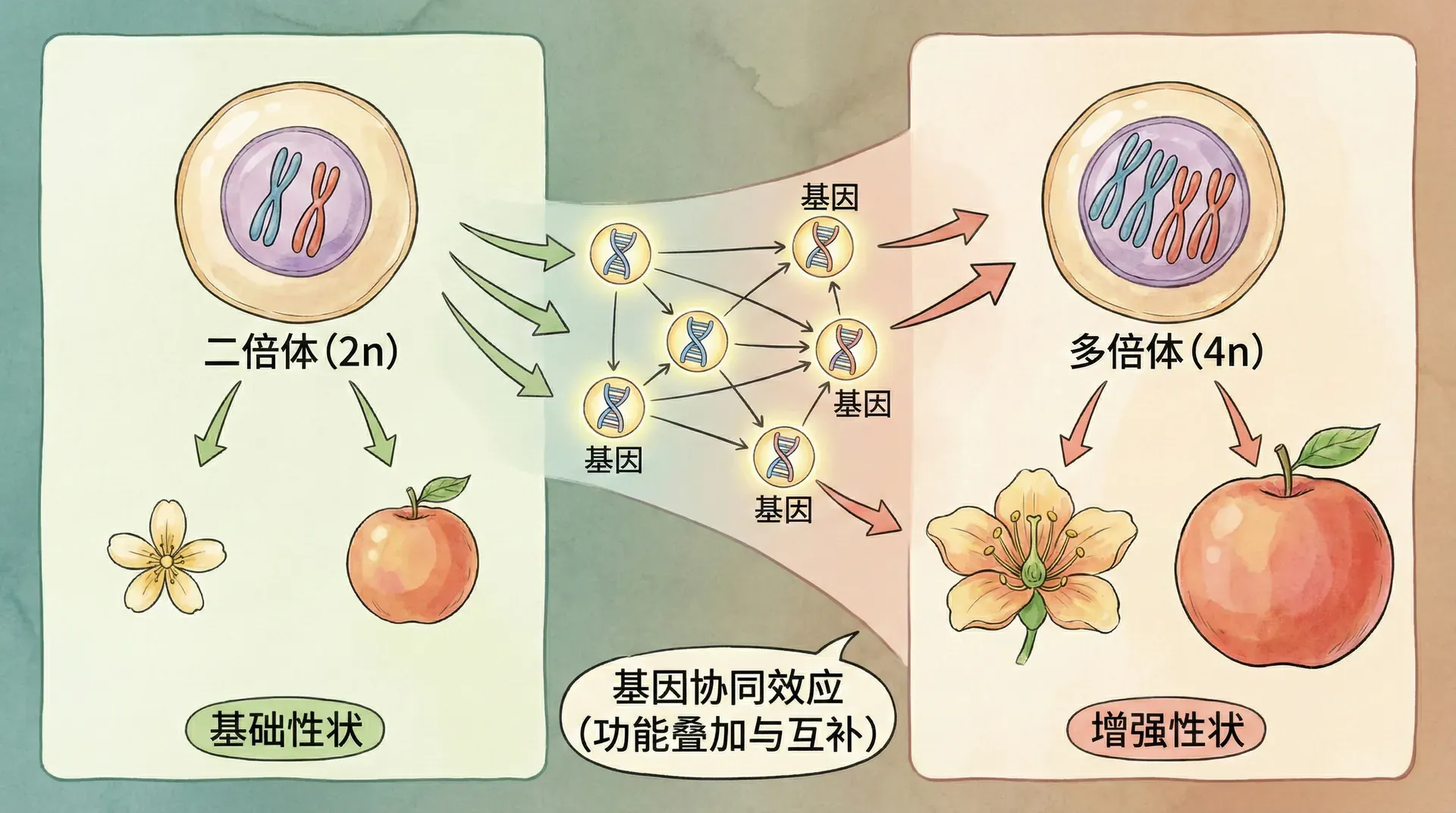
除了常见的染色体重组,有时候自然界和实验中还会发生一种特殊现象——染色体组数异常增加,称为“多倍体”。19世纪末,德弗里斯在研究美洲月见草时首次记录了多倍体现象:这些变异植株外形和普通植株明显不同,后来证实它们的细胞内染色体数量比正常多出一倍。
多倍体产生的机制如下所示:
例如,在极少数情况下,受精卵在发育早期复制了全部染色体,但细胞没有立即分裂。等细胞真正分裂时,每个子细胞都拥有比正常多一套完整染色体,这就是二倍体变为四倍体、三倍体等多倍体现象的由来。
你可能会好奇:既然染色体和基因都是成倍增加,为何会诞生全新的性状?比如原本植株有1个红花基因和1个白花基因,现在变成2个红、2个白,难道不会和从前一样吗?
实际上,这背后涉及基因间的协同作用。基因不会“各自为政”,它们之间常常通过复杂的网络共同影响性状表现。可以将其比作一个交响乐团——每个基因就像一种乐器,单独演奏时旋律有限,但当乐器数量翻倍,即便没有新乐谱,整体音乐效果也会发生巨大的层次变化。正因为如此,基因数目或染色体组数的变化,往往带来意想不到的新性状。
多倍体在农业上的应用
中国是农业大国,多倍体技术已广泛用于作物和动植物的育种。例如:
此外,许多粮食作物(如小麦、甘蔗、棉花),以及一些观赏植物和蔬果,也大量应用多倍体选育技术,极大丰富了品种和经济价值。
多倍体现象在植物界尤为常见,对农业育种意义重大。我国诸多优良作物与园艺品种都离不开多倍体技术的应用。
不仅如此,即使不改变染色体总数,染色体片段的重排(如倒位、易位)也会对性状产生影响。在交叉互换过程中,染色体片段有时会被插到新的位置或发生方向颠倒。这就像房间里的家具全部还在,但重新摆放后整体氛围发生了变化,基因的相对位置和相互作用因此改变,生物也可能呈现出新的特征。
因此,多倍体和染色体结构重排共同推动了生物多样性的产生,是遗传变异和生物进化的重要动力之一。
总结
我们讨论了染色体重组、交叉互换、多倍体、染色体片段重排等多种产生遗传变异的方式。但从长远来看,最重要的突变来源并不是这些——而是全新等位基因的产生。
之前提到的所有机制,本质上都是在现有基因的基础上重新排列组合。这就像用同一副扑克牌可以玩出很多种游戏,但无论怎么洗牌,红桃A还是红桃A。真正革命性的变化,来自于基因本身的改变:原有的基因发生化学结构改变,变成了一个全新的等位基因,就像红桃A突然变成了黑桃K。
这种全新等位基因的形成机制,涉及DNA分子层面的变化,是遗传学继续深入研究的方向。可以说,染色体层面的各种重组为生物提供了丰富的遗传多样性,而基因层面的突变则为进化提供了真正的原材料。两者相辅相成,共同造就了地球上生命的多样性和复杂性。
染色体的变化机制让我们理解了遗传多样性的来源。配对、分离、重组、交叉互换,这些过程看似复杂,实则遵循着清晰的规律,既保证了遗传信息的稳定传递,又创造了充足的变异空间,为生命的延续和进化奠定了基础。