细胞的繁衍
当我们看到一个新生儿呱呱坠地、发出生命的第一声啼哭时,很少有人会想到,这个复杂而奇妙的生命体,最初仅仅是由父母各自提供的两个微小细胞——一个精子和一个卵子——结合而成。这两个肉眼无法直接分辨的细胞,在相遇、融合、基因重组后,形成了一个全新的受精卵。这个受精卵作为新生命的起点,开始经历无数次精确且有序的细胞分裂与分化,从最初的一个细胞逐步发育,最终成长为拥有数万亿个细胞、结构高度复杂的人体。
这其中不仅包括皮肤、骨骼、肌肉等器官和组织的形成,更包含了神经、血液、免疫等多种系统的建立。每一个过程中,都有无数微观的生化反应在默默进行,基因信息被一丝不差地传递下去,环境信号精准地调控细胞的行为。正是这些精密的机制,保证了新生命从无到有,顺利孕育和成长。
精子与卵子
显微镜下的发现
十七世纪末期,当显微镜技术刚刚起步时,研究者们就开始好奇:生命究竟是如何开始的?1677年,科学家首次在显微镜下观察到精液中存在着无数微小的、像蝌蚪一样游动的细胞,这就是精子细胞。这些精子细胞拖着长长的尾巴,在液体中活跃地摆动前进。
然而,仅有精子是不够的。直到1827年,科学家才在雌性哺乳动物体内发现了卵细胞的存在。与鸟类和爬行动物能够产下可见的蛋不同,哺乳动物的卵细胞隐藏在体内,因此发现它们花费了更长的时间。
当细胞学说建立之后,科学家们认识到,精子和卵子都是单个细胞——它们被称为生殖细胞或配子。一个完整的生命个体需要精子细胞与卵细胞融合,形成受精卵,然后才能开始发育。
大小悬殊的两个细胞
精子和卵子在外观上有着巨大的差异。精子细胞非常小,甚至比普通的体细胞还要小,它基本上就是一个紧密包裹着遗传物质的细胞核,外加一条用于运动的尾巴。而卵细胞则大得多,直径约为针尖大小,是人体内最大的细胞之一。
为什么会有如此大的差异?让我们用数据来直观地看一下:
从图中可以看出,卵细胞的直径是精子的24倍,是普通体细胞的6倍。这种大小差异并非偶然。卵细胞之所以这么大,主要是因为它储存了大量的营养物质,这些营养将在受精卵开始分裂后,为早期胚胎提供能量,直到胚胎能够从母体获取营养为止。
曾经有很长一段时间,人们争论究竟是精子还是卵子在遗传中起主导作用。精子看起来活力十足,而卵子则是被动的。但是,19世纪中期孟德尔的豌豆实验明确地告诉我们:无论是用A品种的花粉(相当于精子)授粉给B品种的卵细胞,还是反过来,结果都是一样的。这证明了精子和卵子对遗传的贡献是完全平等的。它们虽然大小悬殊,但在遗传信息的传递上,作用是相同的。
从一个细胞到完整生命
胚胎发育的奥秘
当精子与卵子结合形成受精卵后,这个单细胞就开始了一段奇妙的旅程。受精卵会不断地分裂:一个变成两个,两个变成四个,四个变成八个……细胞数量呈指数级增长。
在这个过程中,细胞不仅在数量上增加,还会逐渐分化出不同的类型。早期的胚胎会形成三个基础的细胞层:外胚层、中胚层和内胚层。这三个胚层就像是建筑工地的三种基础材料,最终会发育成不同的器官和组织。外胚层将形成皮肤和神经系统,中胚层会发育成肌肉和骨骼,内胚层则构成消化系统和呼吸系统。
中国的胚胎学研究在20世纪有着重要的贡献。著名生物学家童第周在两栖类动物的胚胎发育研究中取得了突破性成果,他的实验揭示了细胞核与细胞质在发育中的相互作用。贝时璋院士则在细胞重建和无核细胞研究方面做出了开创性工作,为理解细胞分化提供了重要线索。
有一个有趣的现象:人类胚胎在发育过程中会短暂地出现一些“多余”的结构。例如,在胚胎早期会形成类似鱼鳃的结构,但很快就会消失或转变成其他器官(如部分变成耳朵的结构)。还有,我们的胚胎会长出尾巴,但在出生前就退化了。这些现象提示我们,个体的发育过程某种程度上反映了物种的进化历程。
同卵双胞胎是如何形成的?当受精卵分裂成两个细胞后,如果这两个细胞意外地分开了,每个细胞都可以独立发育成一个完整的个体。由于它们来自同一个受精卵,因此拥有完全相同的遗传信息,外貌也几乎一模一样。
细胞分裂的精密机制
既然生命从一个细胞开始,那么细胞是如何分裂的呢?这个过程必须极其精确,因为每个新细胞都需要获得完整的遗传信息。
在细胞内部,有一个被膜包裹的结构叫做细胞核,它是细胞的“指挥中心”。细胞核内含有染色质,这些染色质在细胞分裂时会浓缩成清晰可见的棒状结构,称为染色体。染色体携带着全部的遗传信息。
19世纪后期,随着染色技术的发展,科学家们终于能够清楚地看到细胞分裂过程中染色体的变化。他们发现,细胞分裂并非简单地一分为二,而是经过一系列精心编排的步骤,确保每个子细胞都能获得准确的遗传信息副本。
人体的大多数细胞都含有46条染色体(23对)。这个数字必须在每次细胞分裂后保持不变。那么细胞是如何做到的呢?答案就在于一种称为“有丝分裂”的机制。
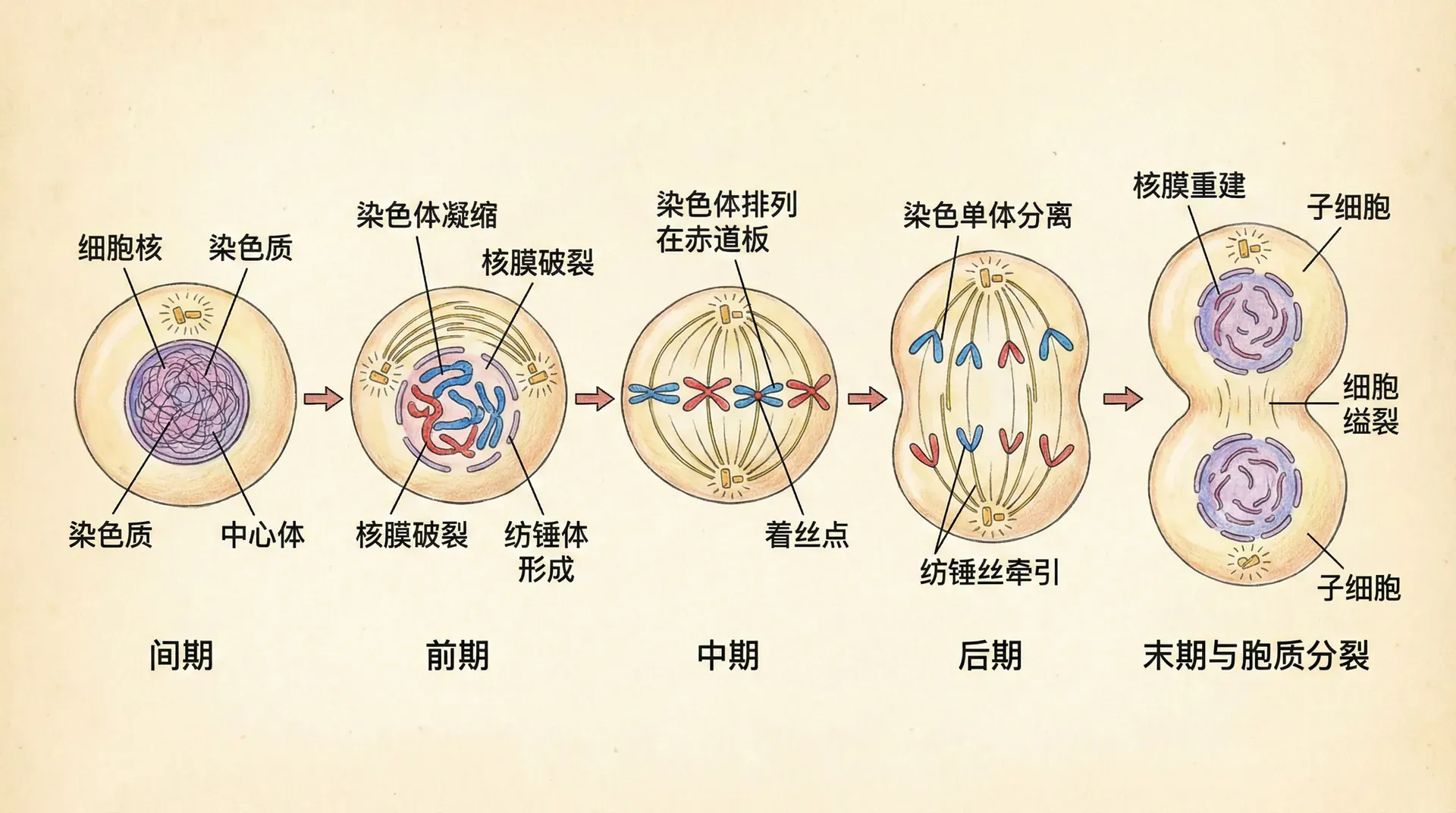
有丝分裂
染色体的舞蹈
有丝分裂是体细胞进行分裂的方式,确保遗传信息准确无误地传递给子代细胞。整个过程可以分为几个连续的阶段,就像一场精心编排的舞蹈。
在细胞准备分裂之前,处于间期的细胞看起来相对平静。细胞核清晰可见,染色质松散地分布在核内。在细胞核旁边,有一个微小的结构叫做中心体,它将在分裂中起到关键作用。
当分裂开始时,首先进入前期。此时,染色质开始凝聚,逐渐形成可见的染色体。中心体一分为二,两个中心体开始向细胞的两极移动。同时,细胞核的核膜开始解体,染色体完全暴露在细胞质中。
接下来是中期。两个中心体已经到达细胞的两端,它们之间伸出许多纤维状的结构(纺锤丝)。染色体排列在细胞的中央,形成一个整齐的平面,就像士兵列队一样。这是观察染色体的最佳时期,因为此时染色体浓缩到最紧密的状态。
然后进入后期。每条染色体沿着纺锤丝被拉向细胞的两极。染色体开始移动,一半向一端,另一半向另一端。细胞的中央区域逐渐变空。
最后是末期。到达两极的染色体开始松散,恢复成染色质状态。新的核膜在两组染色体周围形成。同时,细胞在中央部位开始收缩,最终分裂成两个独立的子细胞。
有丝分裂过程中染色体数量的变化(以人体细胞为例):
从图中可以看出,有丝分裂的关键特点是:染色体数量始终保持不变。那么细胞是如何做到的呢?
有丝分裂的秘密在于:在分裂之前,每条染色体都会复制一份,变成两条完全相同的姊妹染色单体连接在一起。当细胞分裂时,这两条染色单体分开,分别进入两个子细胞。因此,虽然染色体被“分配”了,但由于事先进行了复制,每个子细胞最终仍然拥有完整的46条染色体。
各个阶段的细节
为了更清晰地理解有丝分裂的各个阶段,我们可以将它们的特征整理如下:
有丝分裂在我们的生活中无时无刻不在发生。你的皮肤细胞每天都在进行有丝分裂,替换掉老化或受损的细胞。一个成年人每秒钟大约有2500万个细胞在进行分裂。从婴儿成长到成人,从伤口愈合到头发生长,都离不开有丝分裂的精确运作。
减数分裂
染色体数量减半的意义
现在我们知道了普通体细胞是如何分裂的,但生殖细胞的情况有所不同。科学家们在19世纪末发现了一个令人惊讶的事实:精子和卵子的染色体数量只有体细胞的一半。
以人类为例,我们的体细胞含有46条染色体,但精子和卵子各只有23条。这种现象最初让研究者们感到困惑,但很快就明白了其中的道理。
如果精子和卵子都含有46条染色体,那么受精卵就会有92条染色体。下一代的生殖细胞再结合,就会有184条染色体。这样下去,染色体数量会不断翻倍,显然是不合理的。
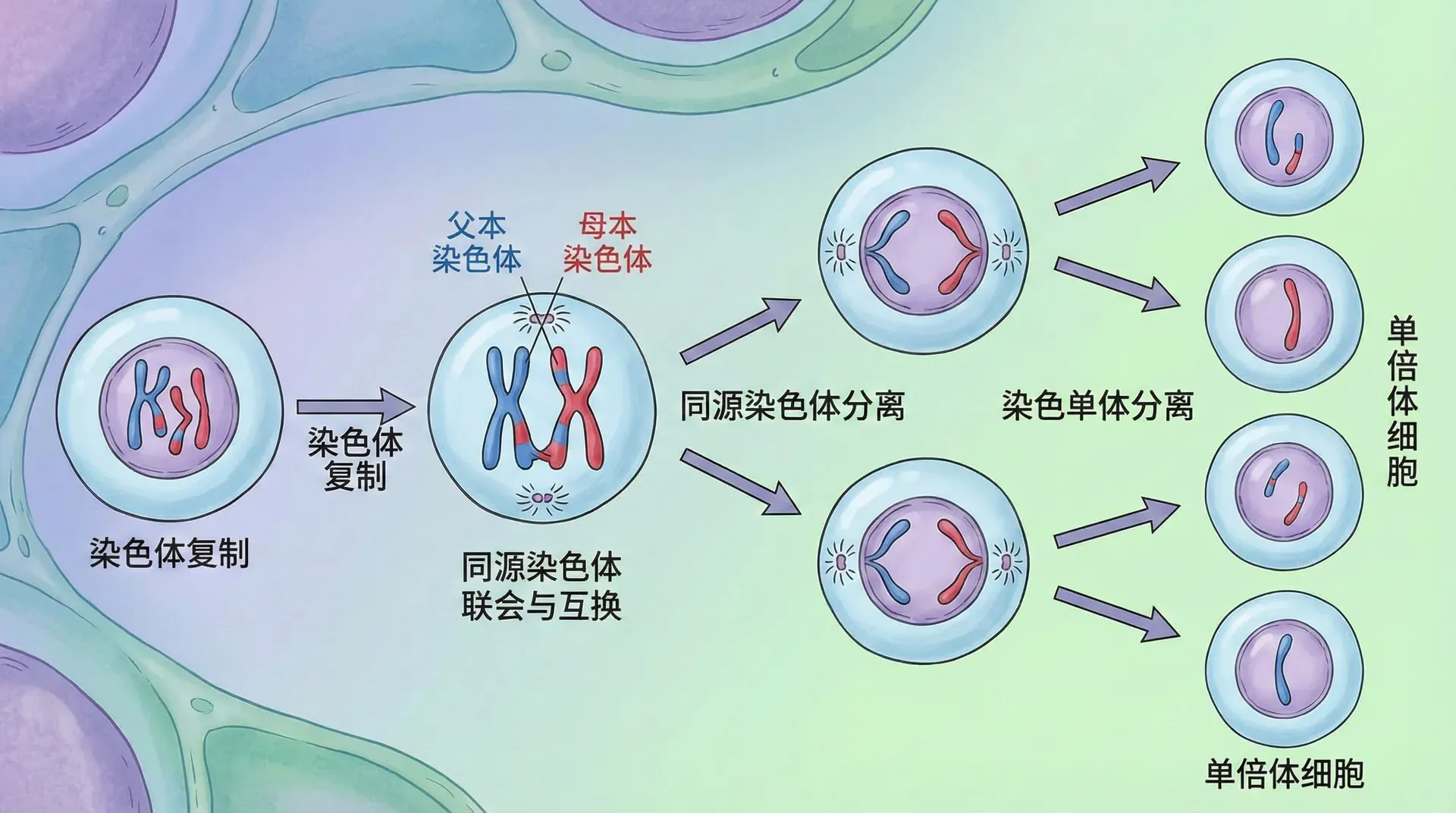
因此,生殖细胞必须通过一种特殊的分裂方式,将染色体数量减半。这种分裂方式叫做减数分裂。我们把染色体数量正常的细胞称为二倍体(用2n表示),而染色体数量减半的细胞称为单倍体(用n表示)。
下方是减数分裂中染色体数量的变化:
从图中可以看到,减数分裂经历了两次连续的细胞分裂,但染色体只复制一次。第一次分裂(减数分裂I)使染色体数量减半,从46条变成23条。第二次分裂(减数分裂II)类似于有丝分裂,保持染色体数量不变,但细胞数量再次加倍。最终,一个初始细胞产生四个配子,每个配子含有23条染色体。
当精子(23条染色体)与卵子(23条染色体)结合时,受精卵又恢复到46条染色体,形成一个完整的二倍体细胞。这样,染色体数量就在世代之间保持稳定了。
不同生物的染色体数量差异很大。水稻有24条染色体(12对),大熊猫有42条染色体(21对),而人类有46条染色体(23对)。有趣的是,染色体数量的多少与生物的复杂程度并没有直接关系,一些植物的染色体数量甚至比人类还要多。
遗传物质的随机组合
减数分裂不仅仅是简单地将染色体数量减半,它还有一个重要的作用:增加遗传多样性。
我们前面提到,人体细胞有46条染色体,它们是成对存在的:23对染色体。每一对中,一条来自父亲,一条来自母亲。比如,第1号染色体有两条,一条是父亲的,一条是母亲的;第2号染色体也有两条,以此类推。
在减数分裂时,每一对染色体会分开,但是分到哪个配子中,完全是随机的。以一个简化的例子来说明:假设某生物只有3对染色体,我们可以用A和A'表示第一对,B和B'表示第二对,C和C'表示第三对(其中带撇号的来自父亲,不带撇号的来自母亲)。
当这个生物产生配子时,可能得到的组合有:ABC、ABC'、AB'C、AB'C'、A'BC、A'BC'、A'B'C、A'B'C',共8种不同的组合。而人类有23对染色体,可能的组合数量是2的23次方,也就是超过800万种!
这种随机组合极大地增加了遗传的多样性,这也是为什么同一对父母生出的孩子(除了同卵双胞胎)都有不同外貌和性格的原因。
让我们对比一下有丝分裂和减数分裂的主要区别:
染色体与遗传的关系
基因在染色体上的分布
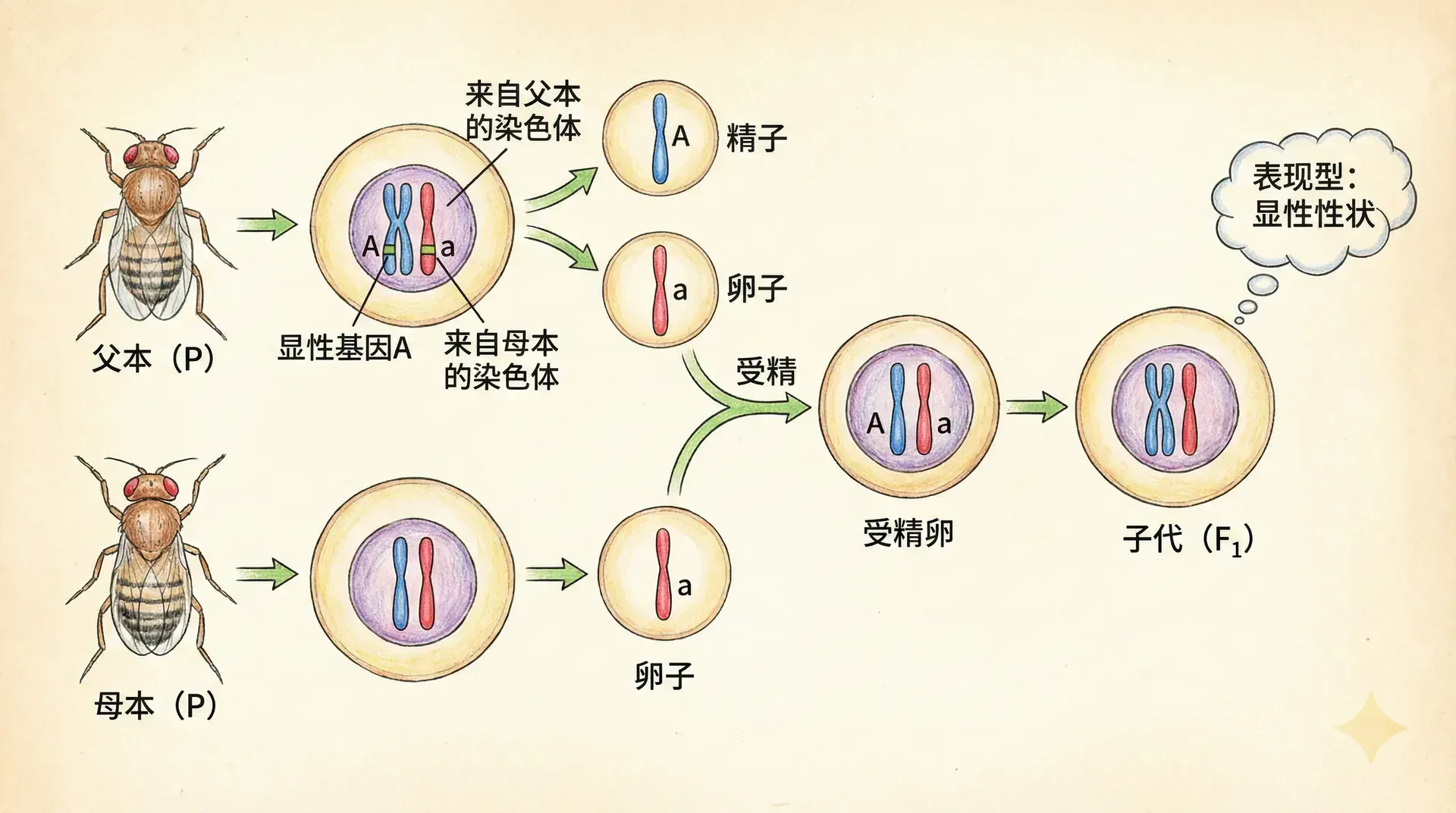
到了20世纪初,两条独立的研究线索汇聚到了一起:一条是孟德尔关于遗传因子的理论,另一条是关于染色体行为的观察。科学家们开始意识到,孟德尔所说的“遗传因子”(现在我们称之为基因)很可能就位于染色体上。
1902年,美国生物学家萨顿明确提出:染色体携带着遗传信息。他注意到,染色体的行为与孟德尔遗传因子的行为惊人地相似。
让我们回顾一下孟德尔的经典实验:当他用红花豌豆与白花豌豆杂交时,第一代全是红花,第二代出现了红花和白花的分离,比例大约是3:1。孟德尔解释说,每个植株有两个遗传因子,一个来自父本,一个来自母本。
假设控制花色的基因位于某一对染色体上。每个体细胞有这一对染色体,因此有两个花色基因。在形成配子时,这对染色体分离,每个配子只得到其中一条染色体,也就只携带一个花色基因。
萨顿提出的“染色体遗传学说”认为:染色体成对存在,基因也成对存在;染色体在减数分裂时分离,基因也随之分离;不同对的染色体自由组合,不同基因也自由组合。这完美地解释了孟德尔定律。
孟德尔定律的染色体解释
当我们研究豌豆的花色,用R表示红花基因(显性),用W表示白花基因(隐性)。
一个红花豌豆可能有两种基因型:RR(纯合子)或RW(杂合子)。白花豌豆只能是WW。这些基因位于一对同源染色体上。
当豌豆形成配子时,染色体对会分离。如果是RW的植株,它产生的配子中,一半得到带R基因的染色体,一半得到带W基因的染色体。因此,配子中R和W的比例是1:1。
现在让我们看看杂交的结果。假设父本和母本都是RW,它们各自产生两种配子:一半含R,一半含W。当这些配子随机结合时,会产生什么结果呢?
从图中可以看出,后代的基因型比例是RR:RW:WW = 1:2:1。由于R是显性基因,RR和RW都表现为红花,只有WW表现为白花。因此,红花:白花 = 3:1,这正是孟德尔观察到的比例。
让我们用一个表格来展示这个过程:
这个表格清楚地展示了为什么后代中红花与白花的比例是3:1。四个格子中,三个表现为红花,一个表现为白花。
染色体理论不仅解释了单个性状的遗传,还解释了多个性状的自由组合。如果两个基因位于不同的染色体对上(比如花色基因在第1对染色体上,种子形状基因在第2对染色体上),那么在减数分裂时,这两对染色体会独立地分配到配子中。第1对染色体分离的结果不影响第2对染色体如何分离,反之亦然。这就解释了孟德尔的自由组合定律。
精子和卵子虽然大小相差悬殊,但它们对遗传的贡献是完全平等的。精子的大小优势在于便于运动,而卵子的大小优势在于提供营养。但在遗传信息的传递上,每个配子都携带23条染色体,各占一半,缺一不可。
通过染色体的精密分配机制,生命得以一代代延续。从一个受精卵开始,经过无数次有丝分裂,最终形成一个完整的生命体。而在生命体成熟后,又通过减数分裂产生新的配子,将遗传信息传递给下一代。这个周而复始的过程,既保证了遗传的稳定性,又通过染色体的随机组合创造了无限的可能性,使得每个生命都是独一无二的。
从显微镜下那些微小的细胞,到宏观世界中丰富多彩的生命形式,细胞的繁衍机制将微观与宏观紧密地联系在了一起。理解了细胞如何分裂和传递遗传信息,我们就理解了生命延续的基本奥秘。