减数分裂和受精作用
在前面的学习中,我们已经了解了孟德尔的遗传定律,特别是分离定律和自由组合定律。这些定律揭示了生物性状遗传的基本规律,但你是否想过:这些遗传定律背后的细胞学机制是什么?等位基因为什么会分离?非等位基因又是如何自由组合的?
例如,人体细胞中有46条染色体,这个数目在人类世世代代的繁衍中保持恒定。父亲的体细胞有46条染色体,母亲的体细胞也有46条染色体。如果精子和卵细胞也各有46条染色体,那么受精卵就会有92条染色体,子代的染色体数目就会加倍。但事实并非如此,这是为什么呢?
答案就在于生殖细胞的形成过程——减数分裂。通过减数分裂,生殖细胞中的染色体数目减半,精子和卵细胞各含23条染色体,受精后形成的受精卵恢复到46条染色体,从而维持了染色体数目的恒定。
减数分裂和受精作用不仅解决了染色体数目恒定的问题,更重要的是,它们为遗传定律提供了细胞学基础,为生物的遗传变异提供了物质基础。理解这两个过程,将帮助我们从细胞水平深入理解遗传现象的本质。
减数分裂的概念与意义
减数分裂(meiosis)是一种区别于有丝分裂的特殊细胞分裂方式。它主要发生在进行有性生殖的生物体内,由体细胞或生殖母细胞经过一系列精细调控的分裂过程,最终产生染色体数目减半的生殖细胞(如精子、卵细胞或植物的孢子)。减数分裂的最显著特征是:整个过程中细胞发生两次连续分裂(分别称为减数第一次分裂和减数第二次分裂),但染色体只复制一次。这样一来,产生的子细胞中染色体数目正好为亲本细胞的一半,为维持生物种群染色体数目的稳定和遗传信息的有序传递提供了基础。
减数分裂发生的时期和场所
减数分裂仅发生在有性生殖生物体生成生殖细胞(或孢子)的过程中。在动物体内,减数分裂主要发生在生殖腺内:雄性动物的精巢产生精子,雌性动物的卵巢产生卵细胞。无论是低等动物还是高等动物,只要依赖有性生殖,其生殖细胞的产生都离不开减数分裂。
在被子植物等高等植物体内,减数分裂则主要在花药(产生花粉,花粉母细胞减数分裂形成花粉)和胚珠(产生卵细胞,胚珠内的大孢子母细胞减数分裂形成大孢子)内进行。此外,某些真菌、藻类等也通过减数分裂形成孢子,为后代的遗传物质减半打基础。可以说,减数分裂是各类有性生殖真核生物的基本生命现象之一。
减数分裂的意义
减数分裂在生物学上具有重要的意义:
因此,减数分裂不仅是有性生殖生物维持遗传稳定和变异的根本保障,也是解释遗传规律和研究生物多样性的“桥梁”与“核心”。理解减数分裂的概念和重要意义,是学习细胞遗传、生命演化乃至现代生物科学不可或缺的基础。
减数分裂的过程
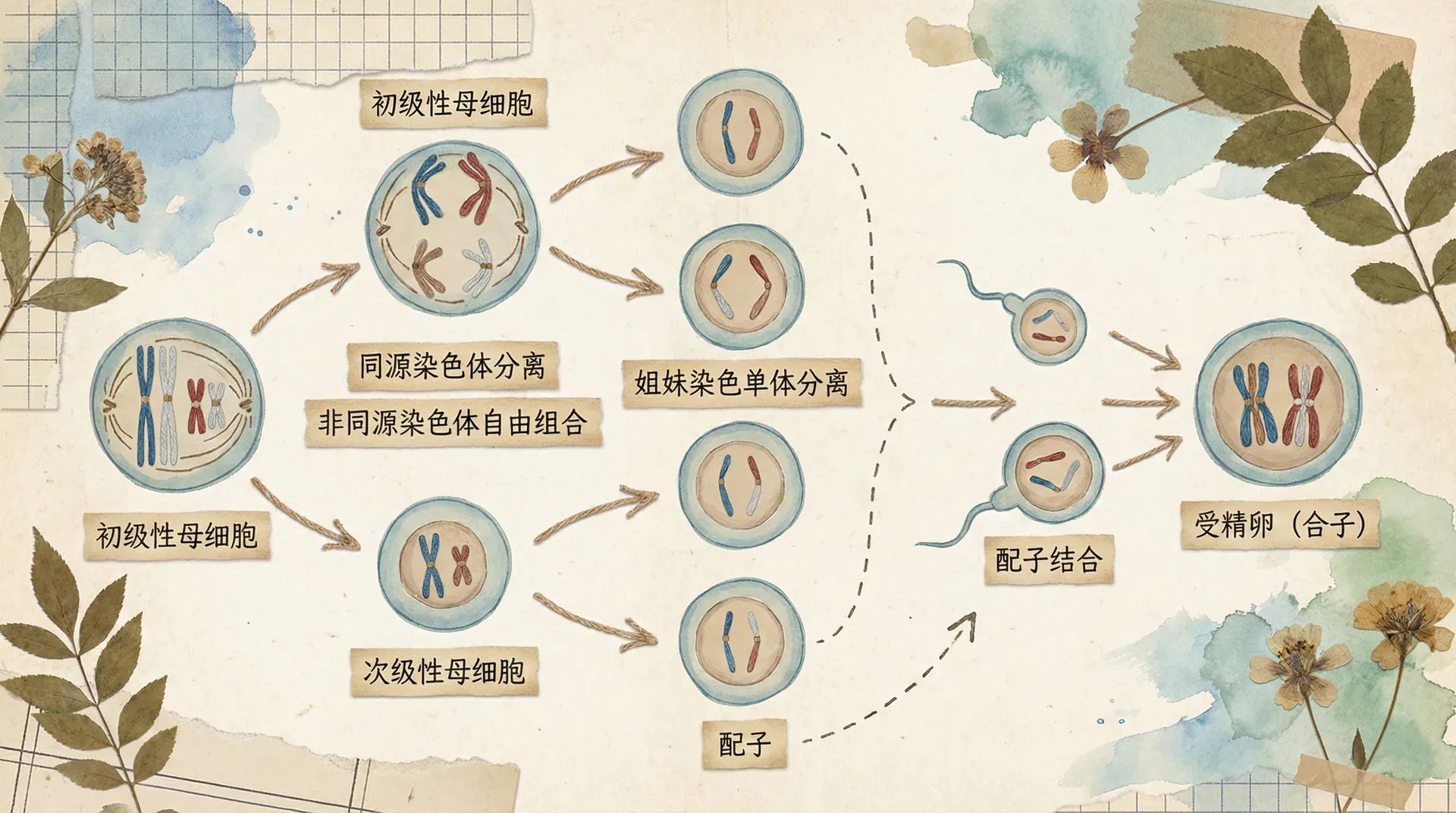
减数分裂包括两次连续的细胞分裂:减数第一次分裂和减数第二次分裂。我们以精子的形成为例,详细了解这个过程。
减数分裂前的间期
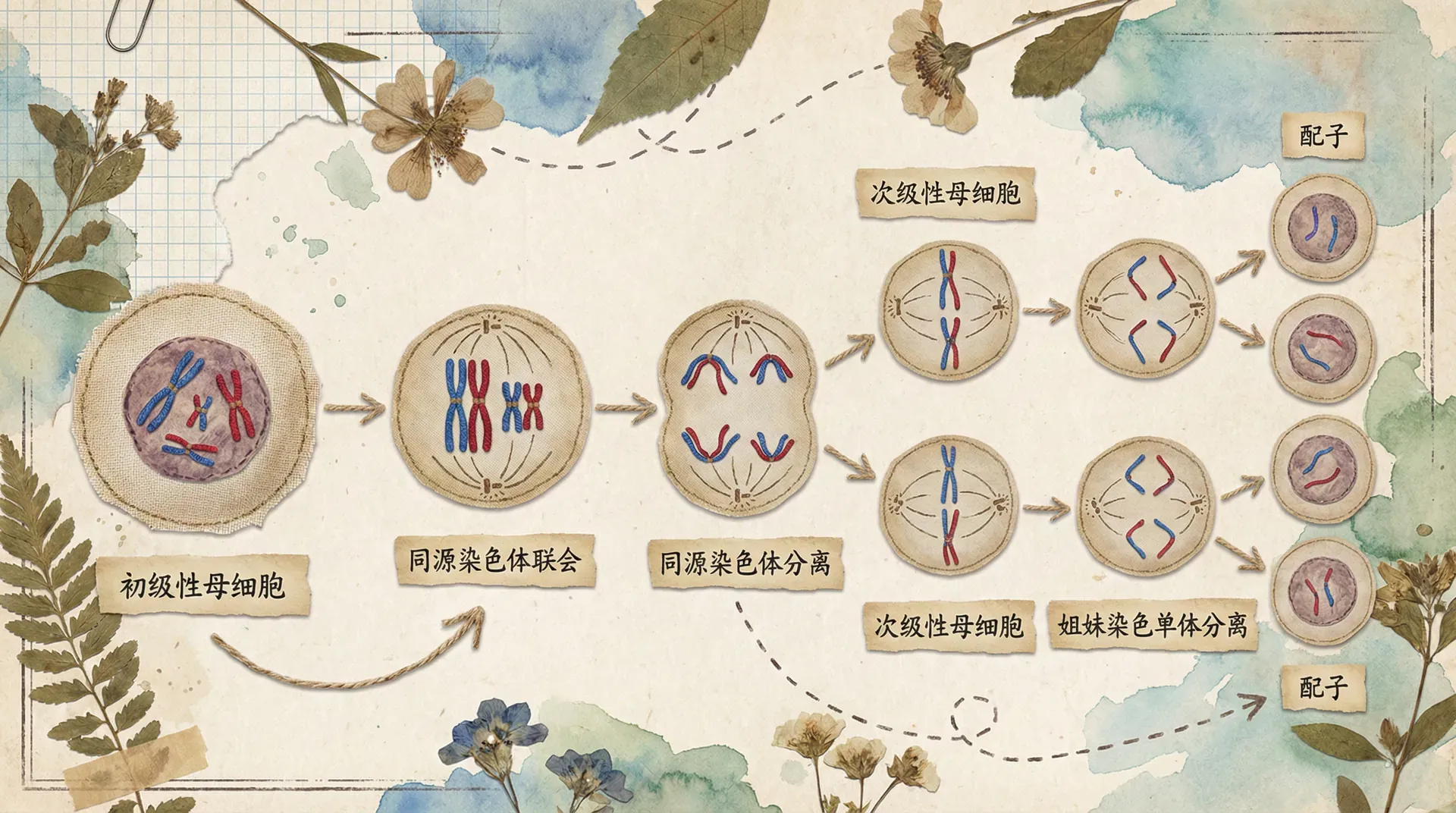
在减数分裂开始之前,有一个间期。这个间期与有丝分裂的间期类似,细胞进行DNA复制,染色体复制形成染色单体,同时合成蛋白质,细胞器增多,为分裂做准备。此时,每条染色体都由两条姐妹染色单体组成,通过着丝点连接在一起。
间期是减数分裂的准备阶段,DNA复制使遗传物质加倍,但染色体数目不变,因为姐妹染色单体共用一个着丝点。
减数第一次分裂
减数第一次分裂是减数分裂中最关键、最特殊的阶段,它包括前期、中期、后期和末期四个时期。
前期是减数分裂中最复杂、历时最长的时期。在这个时期,染色质丝高度螺旋化,缩短变粗,形成染色体。此时最重要的事件是同源染色体联会。同源染色体是指形态、大小相同,一条来自父方,一条来自母方的两条染色体。联会是指同源染色体两两配对,紧密结合在一起的现象。联会后的每对同源染色体称为一个四分体,因为此时每个四分体包含四条染色单体。同时,细胞核膜和核仁逐渐消失,纺锤体开始形成。
中期时,同源染色体成对排列在细胞中央的赤道板两侧。注意,这里是“两侧”而不是“上”,这与有丝分裂中期染色体排列在赤道板上有明显区别。此时,纺锤体完全形成,纺锤丝连接在染色体的着丝点上。
后期是减数分裂的关键时期。在纺锤丝的牵引下,同源染色体彼此分离,分别移向细胞的两极。这个过程非常重要,因为它是孟德尔分离定律的细胞学基础——等位基因随着同源染色体的分离而分离。同时,非同源染色体之间的移向是随机的、自由的,这是自由组合定律的细胞学基础。需要注意的是,此时姐妹染色单体并没有分开,它们仍然通过着丝点连接在一起。
减数第二次分裂
减数第一次分裂结束后,细胞短暂停留,随即进入减数第二次分裂。减数第二次分裂的过程与有丝分裂非常相似,也包括前期、中期、后期和末期四个时期。
前期时,染色体再次浓缩,核膜和核仁消失(如果在末期重新出现的话),纺锤体形成。
中期时,染色体排列在细胞中央的赤道板上。注意这里是“上”而不是“两侧”,与减数第一次分裂中期不同。
后期时,着丝点分裂,姐妹染色单体彼此分离,成为两条独立的染色体,在纺锤丝的牵引下分别移向细胞两极。
末期时,染色体到达两极,核膜和核仁重新出现,纺锤体消失,细胞质分裂,形成两个子细胞。
减数分裂的结果
经过减数第一次分裂和减数第二次分裂,一个精原细胞最终形成四个精细胞。这四个精细胞中的染色体数目是原来的一半,DNA含量也是原来的一半。精细胞经过变形,形成成熟的精子。
以下是减数分裂过程中染色体数目和DNA含量的变化:
从图表中可以清楚地看到,间期DNA复制后,DNA含量加倍(从2变为4),但染色体数目不变(仍为2n)。减数第一次分裂后期,同源染色体分离,染色体数目减半(从2n变为n),DNA含量也减半(从4变为2)。减数第二次分裂后期,着丝点分裂,染色体数目暂时加倍(从n变为2n),但随即在末期Ⅱ分配到两个细胞中,每个细胞的染色体数目为n,DNA含量为1。
精子和卵细胞的形成
虽然精子和卵细胞都是通过减数分裂形成的,但它们的形成过程存在显著差异。
精子的形成过程
精子的形成发生在雄性动物的精巢(睾丸)中。精原细胞通过有丝分裂增殖,部分精原细胞停止增殖,体积增大,成为初级精母细胞。初级精母细胞进行减数第一次分裂,形成两个次级精母细胞。次级精母细胞再进行减数第二次分裂,每个次级精母细胞形成两个精细胞。
因此,一个精原细胞经过减数分裂可以形成四个精细胞。精细胞还需要经过复杂的变形过程才能成为成熟的精子,这个过程包括细胞核浓缩、形成顶体、线粒体聚集形成线粒体鞘、形成尾部等。
卵细胞的形成过程
卵细胞的形成发生在雌性动物的卵巢中。卵原细胞通过有丝分裂增殖,部分卵原细胞停止增殖,体积增大,成为初级卵母细胞。初级卵母细胞进行减数第一次分裂,但这次分裂是不均等的,细胞质的分配极不均等,形成一个较大的次级卵母细胞和一个很小的第一极体。
次级卵母细胞再进行减数第二次分裂,同样是不均等分裂,形成一个大的卵细胞和一个小的第二极体。第一极体也可能进行减数第二次分裂,形成两个第二极体。因此,一个卵原细胞经过减数分裂只形成一个卵细胞和三个极体,极体最终会退化消失。
精子和卵细胞形成过程的比较
精子和卵细胞形成过程的差异主要体现在以下几个方面:
这种差异是生物长期进化的结果,反映了精子和卵细胞在生殖过程中的不同功能。精子数量多、体积小、运动能力强,有利于寻找卵细胞;卵细胞数量少、体积大、营养丰富,为受精后的胚胎发育提供物质基础。
减数分裂与有丝分裂的比较
减数分裂和有丝分裂都是真核生物的细胞分裂方式,但它们在许多方面存在显著差异。理解这些差异,有助于我们深入把握两种分裂方式的本质。
分裂次数和结果的比较
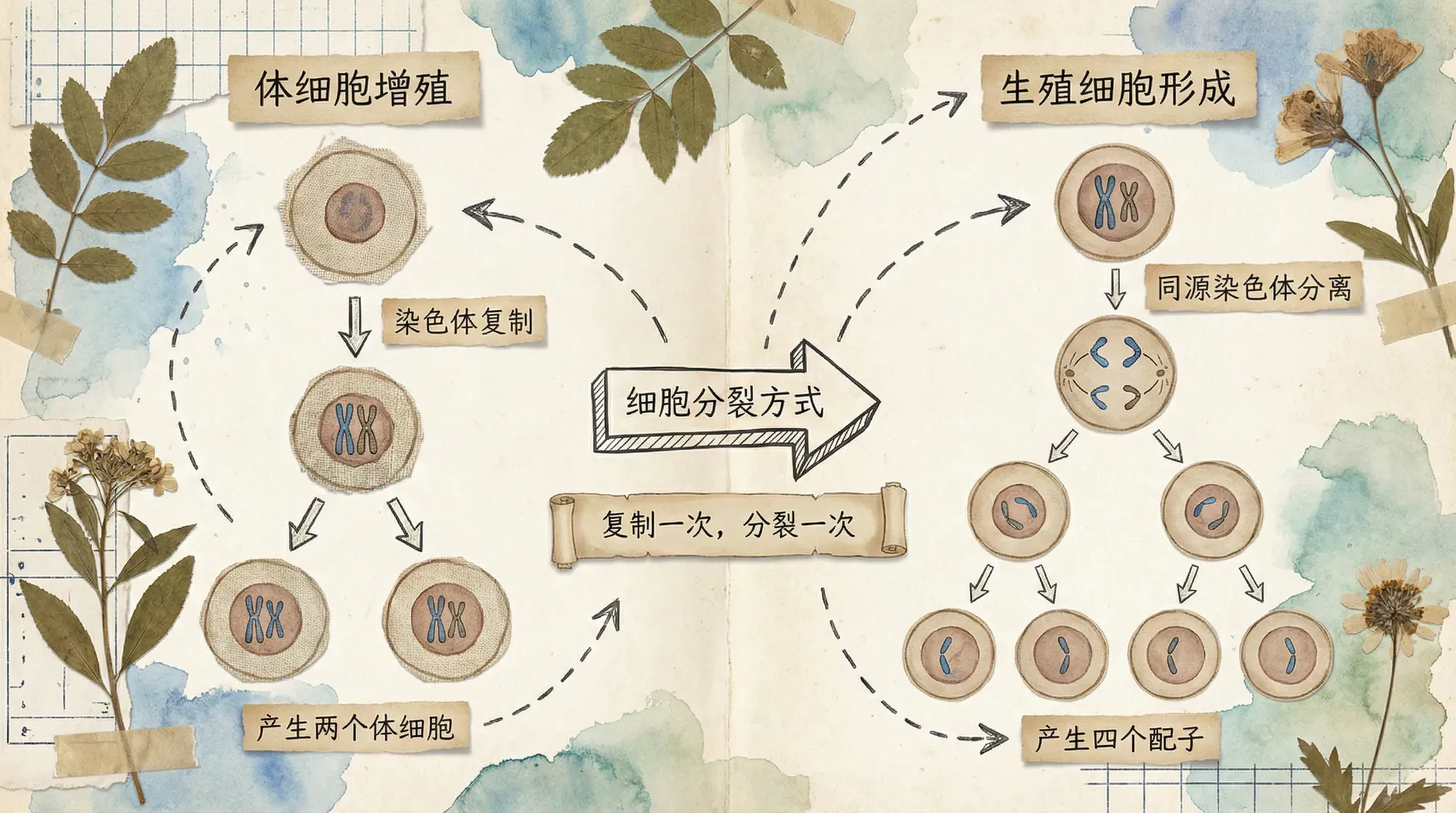
有丝分裂只进行一次分裂,一个母细胞产生两个子细胞,子细胞的染色体数目与母细胞相同。减数分裂连续进行两次分裂,一个母细胞产生四个子细胞(精子形成)或一个子细胞和三个极体(卵细胞形成),子细胞的染色体数目是母细胞的一半。
染色体行为的比较
在有丝分裂中,染色体复制后,姐妹染色单体在后期分离,移向两极,不存在同源染色体的配对和分离。在减数分裂中,减数第一次分裂前期出现同源染色体联会,后期同源染色体分离;减数第二次分裂后期姐妹染色单体分离。
DNA含量和染色体数目变化的比较
下面用图表对比有丝分裂和减数分裂过程中DNA含量的变化:
从图表可以看出,有丝分裂和减数分裂在间期都进行DNA复制,DNA含量从2变为4。有丝分裂一次分裂后,DNA含量减半为2,与原来相同。减数分裂经过两次分裂,第一次分裂后DNA含量为2,第二次分裂后DNA含量为1,是原来的四分之一。
发生范围和意义的比较
减数分裂是有性生殖的细胞学基础,它通过染色体数目的减半和遗传物质的重新组合,既保证了物种遗传的稳定性,又为生物的变异提供了材料。
受精作用
减数分裂使生殖细胞中的染色体数目减半,那么如何恢复到原来的数目呢?答案就是受精作用。
受精作用是指精子和卵细胞相互识别、融合,形成受精卵的过程。这是有性生殖中最关键的环节,标志着新生命的开始。
受精作用的过程
受精作用是一个复杂而精确的过程。首先,精子要穿过卵细胞外围的保护层,到达卵细胞表面。精子的顶体释放出顶体酶,溶解卵细胞的外层,使精子能够进入卵细胞。当一个精子进入卵细胞后,卵细胞会立即发生变化,形成受精膜,阻止其他精子进入,这个机制保证了正常受精,避免多精入卵。
精子进入卵细胞后,精子的细胞核与卵细胞的细胞核融合,形成受精卵的细胞核。这个过程中,来自父方的染色体和来自母方的染色体结合在一起。如果精子和卵细胞各含n条染色体,受精卵就含有2n条染色体,恢复了体细胞的染色体数目。
受精作用的意义
受精作用具有重要的生物学意义。第一,受精作用使精子和卵细胞的染色体结合在一起,恢复了本物种体细胞的染色体数目,维持了物种染色体数目的恒定。第二,受精作用使来自父母双方的遗传物质结合在一起,形成了新的基因组合,这是子代具有双亲遗传特性的根本原因,也是生物多样性的重要来源。
以下是示受精作用前后染色体数目的变化:
从图表可以清楚地看到,人体细胞有46条染色体,经过减数分裂形成的精子和卵细胞各有23条染色体,受精作用后受精卵恢复到46条染色体,新个体发育后体细胞仍保持46条染色体。这个循环周而复始,保证了人类染色体数目的世代恒定。
减数分裂和受精作用
分离定律的细胞学基础
在减数第一次分裂后期,同源染色体彼此分离,分别移向细胞两极。由于等位基因位于同源染色体上,所以等位基因也随着同源染色体的分离而分离,分别进入不同的配子中。这就是分离定律的细胞学基础。
例如,对于豌豆的高茎(D)和矮茎(d)这对相对性状,杂合子(Dd)的植株在形成配子时,减数第一次分裂后期,携带D基因的染色体和携带d基因的染色体分离,分别进入不同的配子,形成D配子和d配子,比例为1:1。
自由组合定律的细胞学基础
在减数第一次分裂后期,非同源染色体的分离是相互独立的、随机的。位于非同源染色体上的非等位基因,在形成配子时可以自由组合。这就是自由组合定律的细胞学基础。
例如,对于豌豆的高茎(D)和矮茎(d)、黄色(Y)和绿色(y)两对相对性状,如果D/d基因位于一对同源染色体上,Y/y基因位于另一对同源染色体上,那么双杂合子(DdYy)在形成配子时,D可以与Y组合,也可以与y组合;d可以与Y组合,也可以与y组合,形成DY、Dy、dY、dy四种配子,比例为1:1:1:1。
受精作用与遗传物质的传递
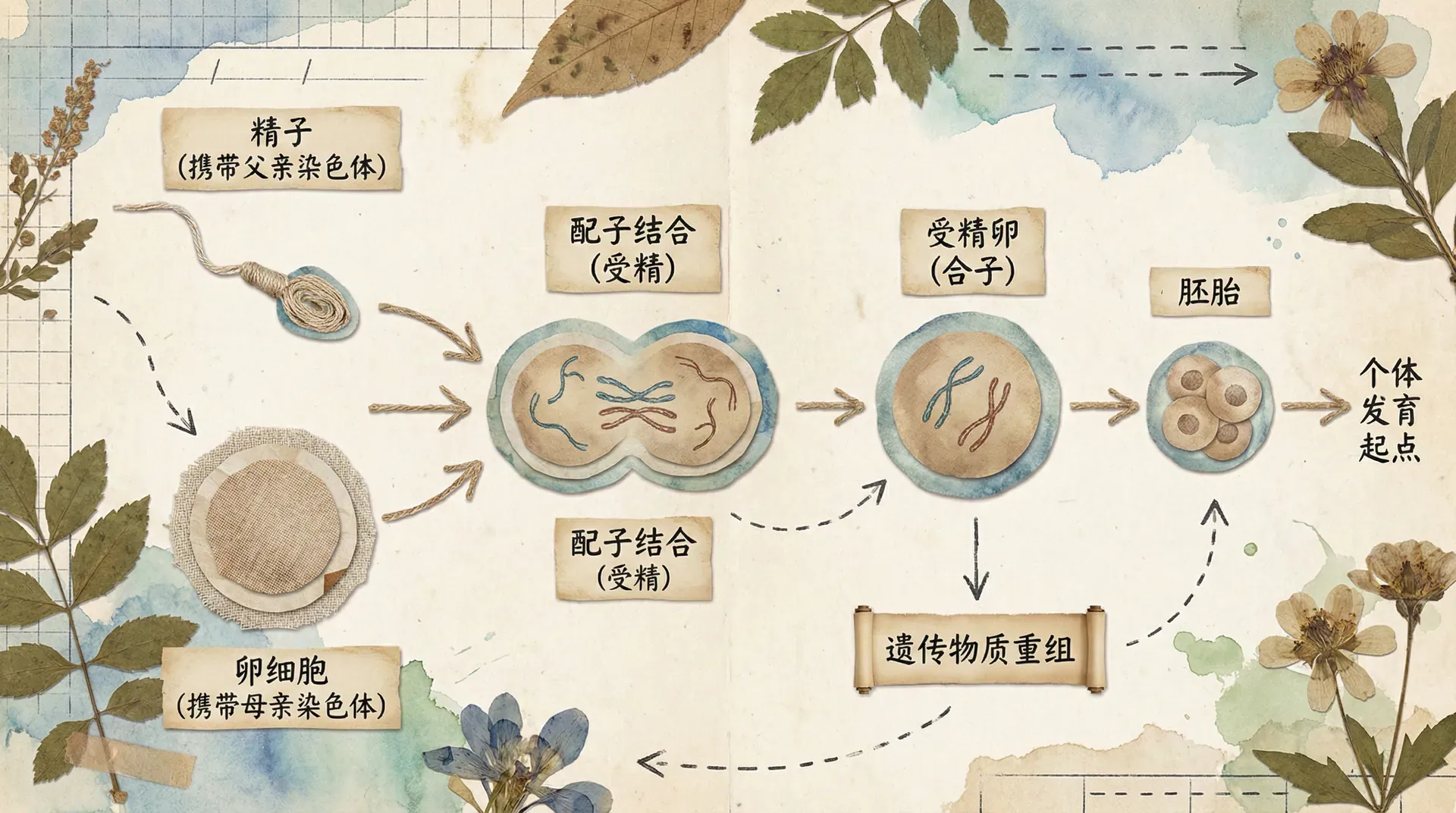
受精作用使来自父方和母方的配子融合,形成受精卵。受精卵中既有来自父方的遗传物质,也有来自母方的遗传物质,这就是子代同时具有父母双方遗传特性的原因。
受精过程中,不同类型的精子与不同类型的卵细胞随机结合,产生了多种多样的基因组合,这是生物多样性的重要来源。例如,人类的精子有2²³种可能的染色体组合,卵细胞也有2²³种可能的染色体组合,理论上一对夫妇可以产生2²³×2²³=2⁴⁶种不同的后代,这个数字超过70万亿,这就解释了为什么除了同卵双胞胎外,每个人都是独一无二的。
需要注意的是,自由组合定律只适用于位于非同源染色体上的非等位基因。如果两对基因位于同一对同源染色体上,它们的遗传就不遵循自由组合定律,而是表现出连锁遗传的现象。
减数分裂和受精作用的意义
减数分裂和受精作用是有性生殖的两个核心环节,它们共同完成了遗传物质的传递和重组,具有重要的生物学意义。
维持染色体数目的恒定
减数分裂使生殖细胞的染色体数目减半,受精作用使受精卵的染色体数目恢复,这两个过程相互配合,保证了生物前后代体细胞染色体数目的恒定。这是物种稳定性的基础,也是生物能够世代延续的前提。
产生遗传变异
减数分裂过程中,同源染色体的分离和非同源染色体的自由组合,产生了多种多样的配子类型。受精过程中,不同配子的随机结合,又产生了更多的基因组合。这些过程为生物的遗传变异提供了丰富的材料,是生物多样性的重要来源,也是生物进化的基础。
实现遗传信息的传递
通过减数分裂和受精作用,遗传信息从亲代传递给子代。子代既继承了父方的遗传信息,也继承了母方的遗传信息,形成了新的基因组合。这种遗传信息的传递和重组,既保证了物种的延续,又产生了个体的差异。
总结
减数分裂是一种特殊的细胞分裂方式,它包括两次连续的分裂,使生殖细胞的染色体数目减半。减数第一次分裂中,同源染色体联会、分离,这是分离定律的细胞学基础;非同源染色体自由组合,这是自由组合定律的细胞学基础。减数第二次分裂类似于有丝分裂,姐妹染色单体分离。
精子和卵细胞的形成过程都经历减数分裂,但存在显著差异。精子形成过程是均等分裂,一个精原细胞产生四个精子;卵细胞形成过程是不均等分裂,一个卵原细胞只产生一个卵细胞和三个极体。
受精作用是精子和卵细胞融合形成受精卵的过程,它使染色体数目恢复,并产生新的基因组合。减数分裂和受精作用共同维持了染色体数目的恒定,为生物的遗传变异提供了材料,实现了遗传信息的传递。理解减数分裂和受精作用,不仅帮助我们深入理解遗传定律,也为我们学习遗传变异、生物进化等内容奠定了基础。
本节练习
第一题:下列关于减数分裂的叙述,正确的是( )
A. 减数分裂过程中,染色体复制两次,细胞分裂两次
B. 减数分裂过程中,同源染色体分离发生在减数第二次分裂
C. 减数分裂形成的生殖细胞中,染色体数目是体细胞的一半
D. 减数分裂只发生在植物的花粉和胚珠中
答案:C
解析:减数分裂过程中,染色体只复制一次(在间期),细胞分裂两次,所以A错误。同源染色体分离发生在减数第一次分裂后期,不是减数第二次分裂,所以B错误。减数分裂形成的生殖细胞中,染色体数目确实是体细胞的一半,C正确。减数分裂不仅发生在植物中,也发生在动物的生殖腺中,所以D错误。本题考查减数分裂的基本特征,重点是理解"一次复制,两次分裂"的特点。
第二题:人的精原细胞中有46条染色体,那么在减数第一次分裂后期,移向细胞一极的染色体数目是( )
A. 23条
B. 46条
C. 92条
D. 无法确定
答案:A
解析:减数第一次分裂后期,同源染色体分离,分别移向细胞两极。人的精原细胞有46条染色体,即23对同源染色体。分离后,移向每一极的是23条染色体(每条染色体仍由两条姐妹染色单体组成)。需要注意区分染色体数目和染色单体数目,此时移向一极的有23条染色体,但有46条染色单体。本题考查减数分裂过程中染色体数目的变化,重点是理解同源染色体分离的含义。
第三题:下表是精子形成和卵细胞形成过程的比较,请将表格补充完整。
答案: ① 精巢(或睾丸) ② 不均等分裂 ③ 4个精子(或一个精原细胞产生4个精子) ④ 保留全部营养物质,为受精卵发育提供物质基础(或类似表述)
解析:本题考查精子和卵细胞形成过程的比较。精子形成发生在雄性动物的精巢(睾丸)中,细胞质均等分配,一个精原细胞产生4个精子,数量多有利于提高受精机会。卵细胞形成发生在雌性动物的卵巢中,细胞质不均等分配,一个卵原细胞只产生1个卵细胞和3个极体,卵细胞大、营养丰富,为受精卵发育提供物质基础。理解这些差异的生物学意义是本题的关键。
第四题:某动物的体细胞中有4条染色体,分别用A、a、B、b表示(A和a是一对同源染色体,B和b是另一对同源染色体)。在减数分裂形成配子时,一个初级精母细胞可以产生几种类型的精细胞?这些精细胞的染色体组成分别是什么?
答案:一个初级精母细胞可以产生2种类型的精细胞。
解析:在减数第一次分裂后期,同源染色体A和a分离,B和b分离。由于一个初级精母细胞中,A和a已经确定,B和b也已经确定,所以分离的结果是确定的。假设A和B移向一极,a和b移向另一极,那么形成的两个次级精母细胞的染色体组成分别是AB和ab。每个次级精母细胞再经过减数第二次分裂,产生两个染色体组成相同的精细胞。因此,一个初级精母细胞最终产生4个精细胞,但只有2种类型:AB型和ab型,各2个。
需要注意的是,如果问的是一个精原细胞可以产生几种类型的配子,答案就不同了。因为在减数第一次分裂时,非同源染色体的分离是随机的,A可以和B一起移向一极(产生AB和ab型配子),也可以和b一起移向一极(产生Ab和aB型配子),所以一个精原细胞理论上可以产生4种类型的配子。但本题问的是一个初级精母细胞,此时染色体的分离方向已经确定,所以只能产生2种类型。
本题考查减数分裂过程中配子类型的判断,重点是理解同源染色体分离和非同源染色体自由组合的含义,以及区分精原细胞和初级精母细胞。
第五题:豌豆的高茎(D)对矮茎(d)为显性,黄色(Y)对绿色(y)为显性,这两对基因分别位于两对同源染色体上。现有一株基因型为DdYy的豌豆,请回答:
(1) 这株豌豆在形成配子时,等位基因D和d的分离发生在减数分裂的哪个时期?
(2) 这株豌豆能产生几种类型的配子?各种配子的比例是多少?
(3) 这株豌豆自交,后代的表型比例是多少?这个比例体现了什么遗传定律?
答案:
(1) 等位基因D和d的分离发生在减数第一次分裂后期。因为等位基因位于一对同源染色体上,当减数第一次分裂后期同源染色体分离时,等位基因也随之分离。
(2) 这株豌豆能产生4种类型的配子:DY、Dy、dY、dy,比例为1:1:1:1。因为D/d基因和Y/y基因位于两对同源染色体上,在减数分裂时,非同源染色体自由组合,D可以与Y或y组合,d也可以与Y或y组合,产生4种配子,且比例相等。
(3) 这株豌豆自交,后代的表型比例是9(高茎黄色):3(高茎绿色):3(矮茎黄色):1(矮茎绿色)。这个比例体现了基因的自由组合定律。
解析:本题综合考查减数分裂与遗传定律的关系。第(1)问考查分离定律的细胞学基础,关键是理解等位基因随同源染色体的分离而分离。第(2)问考查配子类型的判断,关键是理解非同源染色体的自由组合。第(3)问考查遗传定律的应用,DdYy自交的后代表型比例9:3:3:1是自由组合定律的典型比例。通过本题,我们可以看到减数分裂是遗传定律的细胞学基础,遗传定律是减数分裂的宏观表现。
第六题:下列关于受精作用的叙述,错误的是( )
A. 受精作用是精子和卵细胞融合形成受精卵的过程
B. 受精作用使受精卵的染色体数目恢复到体细胞的水平
C. 受精作用产生了新的基因组合,是生物变异的唯一来源
D. 受精过程中,精子和卵细胞的细胞核融合,形成受精卵的细胞核
答案:C
解析:受精作用确实是精子和卵细胞融合形成受精卵的过程,A正确。受精作用使来自父母双方的染色体结合,受精卵的染色体数目恢复到体细胞的水平,B正确。受精作用产生了新的基因组合,是生物变异的重要来源,但不是唯一来源,基因突变、染色体变异、基因重组等都是生物变异的来源,所以C错误。受精过程中,精子的细胞核与卵细胞的细胞核融合,形成受精卵的细胞核,D正确。本题考查受精作用的概念和意义,重点是理解受精作用在遗传中的作用,以及生物变异的多种来源。
第七题:人类男性体细胞中有22对常染色体和1对性染色体(XY),女性体细胞中有22对常染色体和1对性染色体(XX)。请回答:
(1) 一个男性的精原细胞经过减数分裂,能产生几种类型的精子?(从性染色体角度考虑)
(2) 从理论上讲,一对夫妇生男孩和生女孩的概率各是多少?为什么?
(3) 有人说“生男生女取决于母亲”,这种说法对吗?请说明理由。
答案:
(1) 一个男性的精原细胞经过减数分裂,能产生2种类型的精子:含X染色体的精子和含Y染色体的精子。因为男性的性染色体是XY,在减数第一次分裂后期,X染色体和Y染色体分离,分别进入不同的配子。
(2) 从理论上讲,一对夫妇生男孩和生女孩的概率各是50%。因为男性产生的精子有两种类型(含X和含Y),比例为1:1;女性产生的卵细胞只有一种类型(含X)。受精时,含X的精子与卵细胞结合产生XX(女孩),含Y的精子与卵细胞结合产生XY(男孩),两种情况的概率相等,都是50%。
(3) 这种说法不对。生男生女取决于受精时哪种类型的精子与卵细胞结合。女性只能产生一种类型的卵细胞(含X染色体),而男性能产生两种类型的精子(含X或含Y染色体)。如果含X的精子与卵细胞结合,生女孩;如果含Y的精子与卵细胞结合,生男孩。因此,从遗传学角度讲,生男生女主要取决于男性的精子类型,而不是母亲。
解析:本题考查减数分裂、受精作用与性别决定的关系。人类的性别决定属于XY型,男性是异配性别(产生两种配子),女性是同配性别(产生一种配子)。理解这一点,就能正确回答生男生女的问题。同时,本题也涉及科学素养的培养,纠正“生男生女取决于母亲”的错误观念,树立科学的生育观。
第八题:某生物的体细胞中有8条染色体。请计算:
(1) 一个精原细胞经过减数分裂,理论上可以产生多少种染色体组成不同的精子?(不考虑交叉互换)
(2) 如果考虑受精作用,这种生物的一对亲本理论上可以产生多少种染色体组成不同的后代?
(3) 这说明了减数分裂和受精作用的什么生物学意义?
答案:
(1) 一个精原细胞经过减数分裂,理论上可以产生2⁴=16种染色体组成不同的精子。
解析:体细胞中有8条染色体,即4对同源染色体。在减数分裂时,每对同源染色体的分离是独立的,来自父方的染色体和来自母方的染色体可以随机组合。对于4对同源染色体,每对有2种分离方式,所以总共有2⁴=16种组合方式,即可以产生16种染色体组成不同的精子。
(2) 如果考虑受精作用,这种生物的一对亲本理论上可以产生2⁴×2⁴=2⁸=256种染色体组成不同的后代。
解析:雄性个体可以产生2⁴=16种精子,雌性个体也可以产生2⁴=16种卵细胞。受精时,任何一种精子都可以与任何一种卵细胞结合,所以理论上可以产生16×16=256种染色体组成不同的后代。
(3) 这说明减数分裂和受精作用能够产生大量的遗传变异,为生物的多样性和进化提供了丰富的材料。
解析:通过计算我们可以看到,即使是只有4对染色体的生物,通过减数分裂和受精作用就可以产生256种不同的后代。人类有23对染色体,理论上可以产生2²³×2²³=2⁴⁶种不同的后代,这个数字超过70万亿。这充分说明了减数分裂和受精作用在产生遗传变异方面的巨大作用。
本题考查减数分裂和受精作用中配子类型和后代类型的计算,重点是理解非同源染色体自由组合产生配子多样性的原理。计算公式是:n对同源染色体可以产生2ⁿ种配子,一对亲本可以产生2ⁿ×2ⁿ=2²ⁿ种后代。这个计算不考虑交叉互换,如果考虑交叉互换,变异的可能性会更大。