生命演化中的重大转变
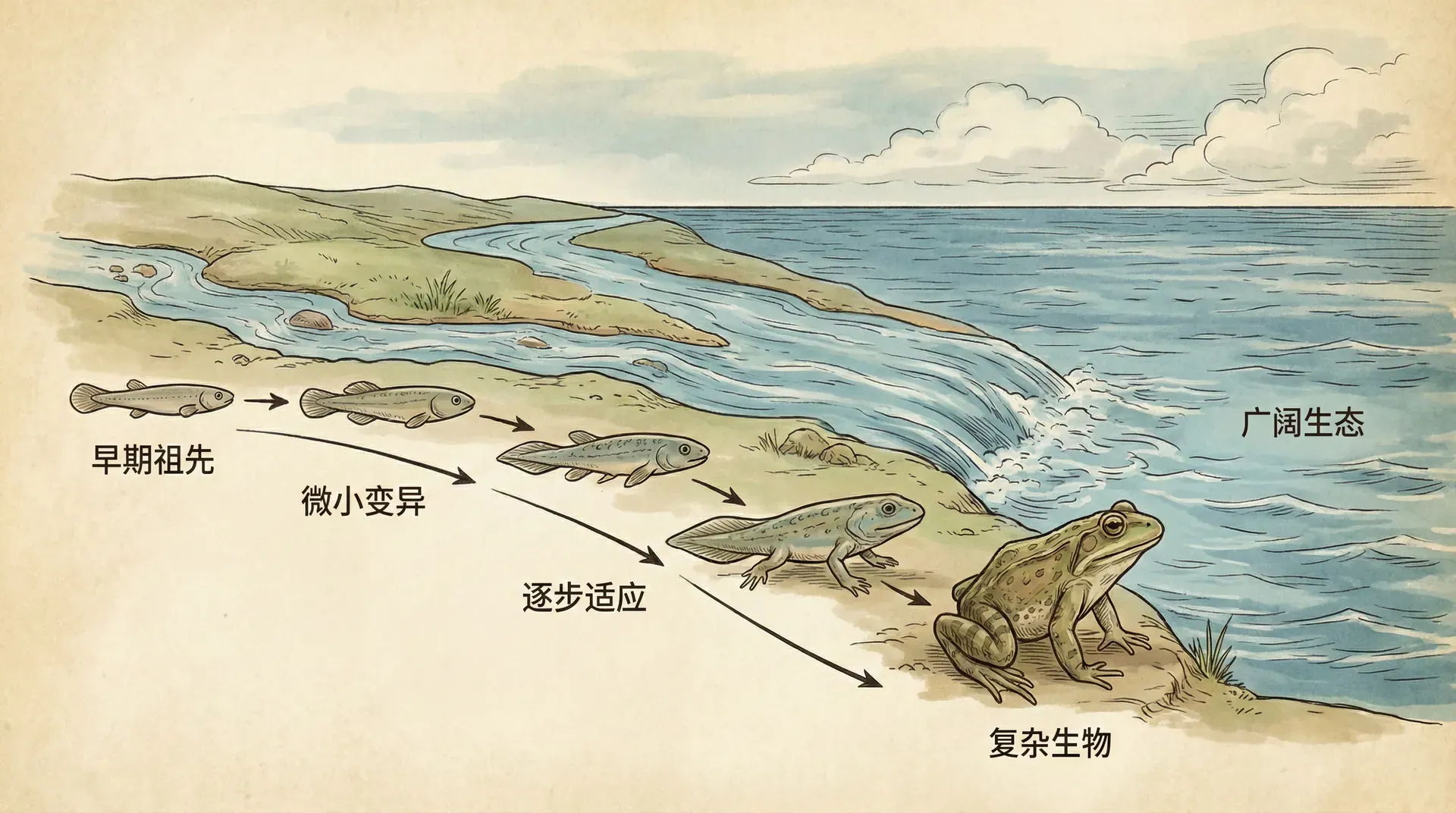
“不积跬步,无以至千里;不积小流,无以成江海。” 生命的演化过程,正如点滴积累汇聚成滔滔江海。每一个微小的变化与关键节点,都推动了生命复杂性的巨大跃升。
地球诞生初期,最早的生命形式必然极其简单,也许只是一些能够自行复制的分子系统。随着时间推移,这些原始生命逐渐经历了难以想象的转变。从单一分子到如今地球上遍布的复杂物种,这一路充满了大量偶然与必然的因素。科学界多年来对这些大尺度变化充满兴趣。越来越多的生物学家认识到,生命历史上的“重大转变”不仅仅是生物体结构和功能上的革命,更关键的是背后普遍存在的适应性挑战。每次转变,实质上都是生物为了解决某种生存难题而作出的创新尝试。
这些创新并非随意发生,而是在演化压力和自然选择的共同作用下,一步步累积并定型。理解这些转变是如何实现的,有助于我们深化对自然选择运作方式的理解,也为揭示现代复杂生物为何会呈现当今的多样面貌提供了强有力的解释。
从简单到复杂的演化历程
在上世纪90年代,英国演化生物学家约翰·梅纳德·史密斯和匈牙利科学家厄尔什·萨特马里,通过对生命历史的深入剖析,系统梳理出一套影响深远的理论体系。他们提出,生命演化史上出现了八个“重大转变”,这些转变的共同特征是——它们极大地改变了遗传信息的传递和复制方式,从而让生命具备了创造新复杂性的能力。每次重大转变,都是遗传层面和协作层面的一次飞跃,让更大的单元能够高效协作并作为整体进行繁衍。
生命演化的八大转变
这八大转变,描绘了一条从最初简单分子进化到人类文明的“生命高速路”:
人类是已知唯一完整经历了所有八次重大转变的生物谱系。这不仅成就了我们个体与群体的复杂性,更决定了我们能够通过文化、教育和技术持续推动自我进化。在“如何演化”这件事上,人类展现了前所未有的多层次和多路径组合。
为什么这些重大转变会反复出现在演化历史中?背后存在一套深刻的挑战与原则:
- 智力层面的挑战:这些转变往往直接影响遗传系统自身,迫使我们从更深入的层级去理解什么是“演化”。基因的“自私”属性、单元间合作与冲突都表现得尤为突出。
- 结构与渐进的挑战:改变之大令人难以置信,比如从单细胞到多细胞,从个体到社会,但每一次转变都必须分解为微小、连续、有益的小步调整,每一步都要保证生物体仍能生存与繁衍。
- 宏观演化层面的挑战:极少数转变会发展成演化上的“制胜法宝”。它们不仅实现了新功能,更被不同谱系在广阔的演化树上不断重复采纳、优化,并延续至今,成为各类复杂生命形式的共同基石。
这些重大转变,是生命演化故事中最为雄浑壮阔的篇章。它们见证了微观分子的聚集协作,也孕育了宏观世界的丰富多样。理解这些转变,有助于我们把握生命的本质及其演化的深层逻辑。
有性生殖的起源
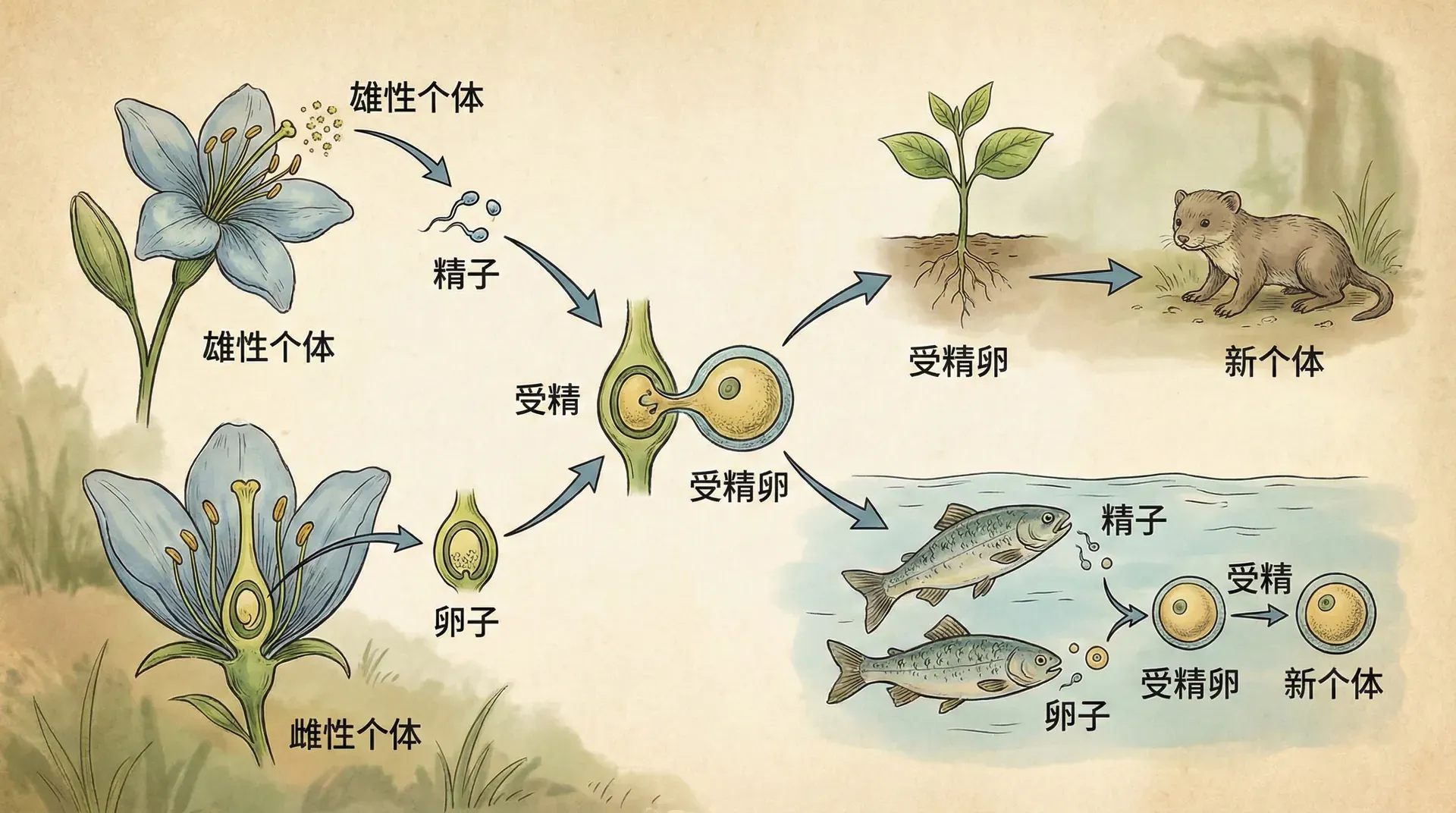
以有性生殖的演化为例,我们可以看到一个生命史中的伟大转折。首先,需要澄清,“性”在生物学语境中,并不仅指交配行为,而是指一种涉及特定细胞周期变化的现象。理解性的演化,关键在于认清细胞如何复制、分裂,以及为何“复制的方式”会对生命系统产生巨大影响。
什么是有性生殖
有性生殖起源于无性繁殖(克隆)祖先,这一过程经过了复杂的进化创新。无性生殖的细胞分裂(有丝分裂)流程较为简单:细胞先复制染色体,然后一分为二。而有性生殖则基于一套更复杂的减数分裂流程:两个单倍体配子(gametes)融合,形成双倍体后代,称为配子融合。配子常常在结构与行为上高度分化——典型如精子(小而能运动)和卵子(大而不动)。此后可能会通过多轮有丝分裂进行发育。在此过程中,还包含一个重要步骤:同源染色体的DNA片段发生交换,即基因重组(交叉互换)。减数分裂会先复制染色体,然后经过两步分裂,产出四个单倍体细胞;部分物种这些单倍体还会再经历有丝分裂,最终再完成配子融合。
以下梳理无性繁殖与有性繁殖主要流程的异同:
有性生殖的演化顺序
有性繁殖并非一蹴而就,而是分阶段逐步形成。部分原生生物(如昆虫肠道中的鞭毛虫)拥有先核内有丝分裂,后减数分裂的周期形态,提示进化的初步为单倍体与双倍体交替周期。此后阶段才发展出配子融合,形成稳定的一步减数分裂(如在孢子虫中所见)。随着演化,交叉互换(基因重组)与染色体加倍机制完善,从而形成两步减数分裂。最终,配子的大小分化演化出现,产生了精子和卵子这样极端分化的配子,这不仅奠定了动物和植物等高级生命的性别基础,也为后续性别相关现象打下基础。
有性生殖重要演化步骤及其生物例子:
基因重组的作用与优势
为什么基因重组是有性繁殖的核心?主要有两个进化动力。
第一,协同突变清除效应。假如突变有协同作用(两个坏突变的损失大于加总值),基因重组能让“坏突变”富集到个别个体上,使其被自然选择迅速剔除,从而降低整个人群的遗传负荷。例如,森林里的橡树、稻田里的水稻,通过基因重组可以更快去除不良突变,获得更强的群体健康度。
为了直观对比有性与无性繁殖种群的遗传负荷分布,请看下图。你会发现有性种群中健康(突变少)的个体更多,突变重灾区则更少——这正是重组带来的效果。
第二动力,则是红皇后假说。多基因性状总被寄生者和宿主之间的进化军备竞赛所推动——就像红皇后(Alice中的角色)飞快奔跑才能原地不动。基因重组的频率和体型、寿命相关,越是长寿、体型大的生物,重组机制越显著,这正好与协同突变和“红皇后压力”假说同时吻合。
基因重组能帮生物清除有害突变,这对于大型、长寿生物尤其重要。比如橡树和大象这种体型大、寿命长的生物,很依赖重组带来的净化效应。现代农作物通过人工控制杂交也正是利用了重组带来的基因优势。
配子的分化与细胞器冲突
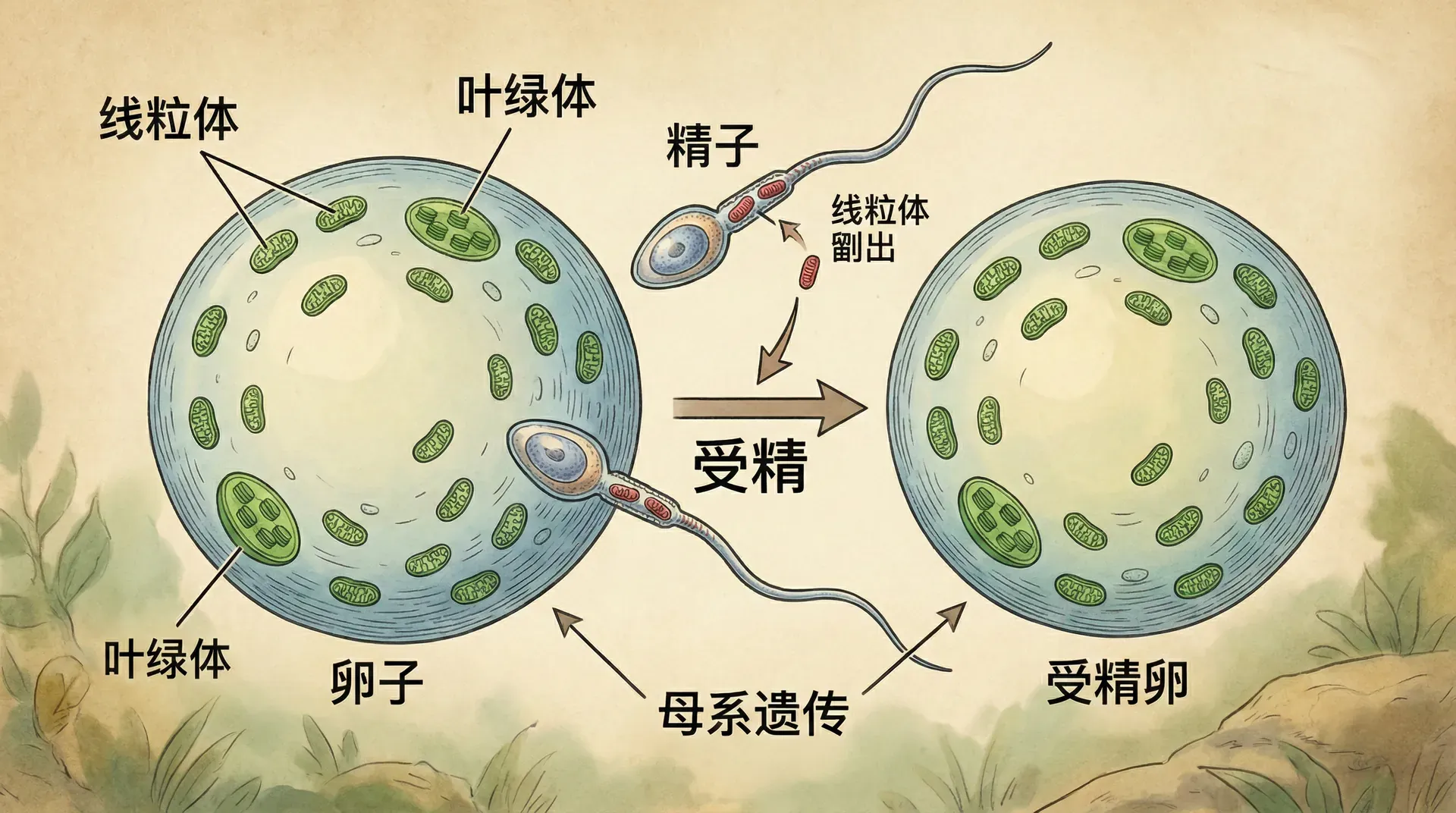
有性生殖最终实现了精子和卵子的深度分化,这一方面源于配子的专职分工,另一方面也关涉细胞器的遗传冲突。许多细胞器如线粒体、叶绿体,都含有自己的基因(它们原本是独立微生物),如果来自两方的细胞器混杂,极易发生竞争从而损伤宿主利益。因此,大多数高级生物采用“单亲细胞器遗传”模式——通常卵子负责传递全部细胞器,精子则负责激活并遗传核DNA。
值得注意的是,这一制度只有两种配子类型时最稳定。如果出现三种或更多配子类型,后代可能会缺失细胞器,造成大问题。
细胞器的单亲遗传只有在交配型不超过两种时才容易维持。细胞器冲突理论提示了两性分化的最深层原因,这也是自然界极少出现多于两性的直接原因。
实际上,部分原生生物(如草履虫、变形虫)通过接合交易遗传信息,却并不交换细胞质,所以也就不存在细胞器争夺。这些特殊类型往往拥有三种、四种甚至更多的不亲和交配型(“类性别”),展示了性别演化的多样性和灵活性。
有性生殖为什么能延续至今
在理解了性的起源和其步步演化的“动力”之后,一个自然的追问是:既然有性生殖存在显著成本,它为何能在绝大多数真核生物中持续和普遍地存在?为什么绝大多数大的谱系都沿袭了有性生殖,而无性繁殖更多见于少数、以“旁枝”形式新近出现的类型?这个问题的解答隐藏着生命多样性和进化的关键机制。
逆转的障碍
有些高度进化的动物和植物类群几乎从未观察到孤雌生殖或长期的无性状态。一个有趣的现象是:虽然性可被轻易丢失,但是逆转回无性却并不易实现。我们可以用表格简明地总结部分群体的“逆转障碍”:
让我们看几个例子:
- 鸟类:虽然偶见孤雌生殖个体(例如家禽中的孤雌出壳小鸡),但因雌性为ZW型(异配性别),孤雌生殖产生卵常含两个Z染色体,生成雄性个体,性别分化得以维持。这也阻碍了单性系的长期建立。
- 哺乳动物:哺乳动物中,基因组印记是逆转障碍的典型。早期实验发现,两个卵的细胞核融合生成的胚胎不能正常发育,因其基因缺少来源差异带来的调控。
- 裸子植物:以松柏为例,卵子一般传递大部分细胞器,但叶绿体只由花粉(雄性)传递。如果只剩单亲繁殖,后代无法获得叶绿体,失去光合作用能力。
此外,在部分动物中(如某些蜥蜴、蚜虫),孤雌生殖通过“依赖精子”模式发生——配子需他种精子刺激才能发育,但精子的遗传物质并不进入后代。这样的机制仍显示了对有性机制残余的依赖。
无性繁殖的短期优势
尽管有上述障碍,无性繁殖一旦成功建立,往往在种群数量增长上表现出巨大的短期优势。很多植物(如蒲公英、甘薯),动物中的部分甲壳类或鱼类,也能长期以无性形式繁衍。
以下信息显示了有性与无性繁殖的子代数量对比(以雌性数量为例):
例如,某些蚜虫种群在夏季几乎完全以无性方式迅速扩张,在同等资源条件下,理论上雌性个体数是有性的两倍。
雌性孤雌生殖者在种群扩展速度上具有决定性的优势——所有后代皆为可生殖个体,不需“浪费”资源产生无繁殖力的雄性。此外,有性生殖还需承担寻找配偶、不匹配等成本。这表面上显示有性个体更易灭绝。
有性生殖的长期优势
奇怪的是,观察现实世界我们会发现大多数无性旁系仅是“新近起源”的例外,大型谱系基本都是长期保有性。例如:
- 鳉鱼(Poecilia)中的鳉鱼和剑尾鱼,有性种起源可追溯数百万年,而孤雌生殖起源则多为几千年内。存活得最久的无性克隆恰恰往往有一定程度的精子表达。
- 水蚤、轮虫、蚜虫等群体,也大多表现为无性旁支短暂繁荣,最终却会被有性谱系“包围”。
这些例外常常与特殊环境、共生关系或其它替代遗传机制(如水平基因转移)相关。
红皇后假说与短期适应
基因重组赋予物种应对迅速变化环境(如寄生虫压力)的能力,红皇后假说就是其中的经典理论。许多实验证据显示,无性种群面临更高的寄生风险。例如轮虫中,无性种群极易被特定病原体灭绝。有性种群凭借遗传变异,可以较快适应新的选择压力。
遗传负荷的累积与穆勒氏棘轮
除了短期适应优势,无性种群面临的最大长期风险是遗传负荷不可逆增加。也就是所谓的“穆勒氏棘轮”效应。其实验和理论解释如下:
- 理论:无性繁殖,基因组以整体方式遗传,无法通过重组恢复无突变型,每发生一次“最优型”的随机丢失就好比棘轮旋转一格,后代无法“倒退”,突变持续累积。
- 实例:轮虫、小型蜥蜴等的无性种,经基因分析发现,基因组中有害突变远高于有性亲缘种,且部分杂合性丧失。
简而言之,有性生殖通过重组和杂合度重建、筛选,有效控制突变累积。而无性种群就像不断“上锁”的棘轮,只能承受越来越高的突变负担,这显著提高了其灭绝风险。
小结
有性生殖能持续存在,归因于:
因此,在进化的大舞台上,有性生殖虽然代价高昂,却以其优势赢得了持久的席位。
总结
有性生殖的演化可以和生命史上的其他重大转变类比,比如细胞起源、染色体形成、从RNA到DNA和蛋白质、真核细胞、多细胞生物、社会群落,以及语言的出现等。这些转变通常都表现为原本相对独立的实体结合成更高层级的整体,有性生殖也是如此:无性个体开始结合,形成有性繁殖种群。
部分演化阶段带来了全新的信息传递方式,比如从RNA到DNA和蛋白质、表观遗传的出现、以及语言让人类获得文化传承。但有性生殖并未引入全新遗传介质,更多是通过重组提升了遗传信息表达与变异的途径。
这些重大转变还有一个共同点——一旦形成就很难逆转。例如细胞结构和有性繁殖的复杂机制一旦建立,都很难退化回原始状态。转变过程中还常见新层级实体间的冲突(如配子来源的细胞器竞争),推动单亲遗传等调控机制,以及功能分工(如雌雄配子差异、社会性协作)等演化策略出现。
需要注意的是,真正意义上的全新信息流系统只出现过三次:分别是DNA-蛋白质体系、表观遗传机制和人类语言。这些极大拓展了生命的复杂性,而有性生殖则着重促进了遗传重组和适应力提升。
综上,研究这些转变帮助我们理解自然选择如何作用于不同谱系层级。每一次突破,都是长期微创新积累与协作的结果,有性生殖的出现也正是复杂生命渐进演化的缩影。