地球生态的演变历程

站在今天中国广袤的土地上,从东部的长江三角洲到西部的青藏高原,从南方的云南热带雨林到北方的大兴安岭针叶林,我们能看到丰富多样的生命形式。昆虫种类繁多,绿色植物覆盖大地,各种脊椎动物和无脊椎动物在不同的生态位中繁衍生息。这些生命几乎遍布地球的每个角落:海洋、淡水、陆地、甚至是缺氧的泥潭、高盐的湖泊、火山温泉、沙漠沙丘,乃至南极冰冻的岩石和地下数百米深的岩层中。
现在地球上最高产、物种最丰富的生态系统都在陆地上。它们包括:绿色植物作为生产者,多样的脊椎动物和昆虫作为植食性动物,捕食者、寄生虫和寄生蜂,还有土壤中的食腐动物、分解者和营养循环细菌。在海洋中,珊瑚礁、海藻林和海草床这些有固着生产者的生态系统最为高产和多样。而开阔的海洋相对贫瘠,主要由浮游植物、浮游动物和大型捕食者组成。生物圈的能量几乎完全来自植物捕获的光能,碳源是大气中的二氧化碳,最终通过需氧呼吸返还给大气。
生物圈的演化历程
生命的起源与早期演化
地球生物圈和生态系统的起源可追溯到距今约38亿至35亿年。关于生命起源,科学家提出了异养起源和自养起源两大理论。早期主流观点认为,最初的生命以原始海洋中合成的有机分子为食,即“异养起源”。但随着对生命化学本质的理解加深,自养理论重新获得重视。原因有两点:一是原始细胞膜通透性有限,难以直接摄取大分子有机物;二是自我强化并可自发出现的代谢循环有可能早于生命出现。若事实如此,生命最早的能量获取模式则应为自养。
最初的生态系统极有可能由化能自养生物构成,而光合作用所需的更复杂机制则在之后逐步发展。伴随化能自养者的出现,很快也会有分解其产物和残骸的第二营养级——这使得生态系统开始分层演化。现有研究也表明,光合作用出现的时间可能比早先认为的更早。值得注意的是,早期生命缺乏蛋白质酶,通常依赖RNA进行催化反应。现代叶绿素的合成过程里,相关分子依然与RNA结合,这被认为是早期RNA催化遗迹。光合作用的出现,为地球系统注入了新的能量流——光能,极大提升了全球生物生产潜力。
早期光合生物可能利用硫化氢或分子氢作为电子供体,而非水。用水作为电子供体时才会释放氧气。
我们已知蓝藻是当今主要产氧原核生物,其历史可追溯至少35亿年。尽管氧气生产持续不断,直到距今约25亿至20亿年,大气中的氧含量才急剧上升,标志着太古代的终结。这期间,必有某些过程大量消耗着产生的氧气。一个关键过程便是需氧呼吸,其产能效率远高于任何厌氧代谢。虽然那时还不存在完整的现代食物链,但需氧呼吸奠定了将来高营养级动物出现的物质基础。至此,碳、氮、硫等主要的生物地球化学循环已基本建立,生命为地球环境注入深刻变革。
真核生物的出现
最早可识别的真核生物化石出现于约21亿年前,大气氧浓度提升阶段。实际上,现代真核生物的祖先谱系非常古老,极可能追溯至最早期(约35亿年前)。在这个阶段中,我们的细菌祖先(生物分子证据显示其与古菌相似)发展出细胞骨架并逐渐丧失了坚硬的细胞壁,具备了吞噬较大型颗粒的能力。最古老的真核生物谱系极可能是厌氧型的。支持这一观点的证据体现在许多现存厌氧真核生物位于真核进化树的早期分支。
真核早期捕食型生物的出现,带来了原核世界中前所未有的营养互动,因为有机物可以被直接摄取,而不是被动通过膜渗透。只有在与线粒体共生、成功整合到需氧环境后,真核捕食者才终于能够高效地利用光合作用所带来的生产力,推动生态系统复杂化。
寒武纪生命大爆发
进入大约10亿年平静期后,地球在5.35亿年前进入剧变。当时的寒武纪大爆发与奥陶纪生物辐射成为进化史的聚光灯。寒武纪期间,主要动物门类轮番登场。我们见证了首批底栖和浮游大型捕食者的产生,也有首批具备穴居能力、能够深入沉积物数厘米的动物。
这种生物多样性的骤增,对地球生态产生了深远影响。生物首次深度操纵了钙、磷、硅等矿物的循环,改变了地质进程。穴居动物的活动促进了营养物质循环,避免了碳和其他元素长期被掩埋。与此同时,新动物们产生大量粪便并沉降海底,有效减少了水体中的溶解氧消耗。地球表层与深层的氧气循环加快,这种变化促进了骨骼化、体型增大及活跃代谢的进化。
澄江生物群(中国云南)展现了寒武纪爆发的壮观景象。5.2亿年前,这些化石记录了数量惊人的生物门类与多样性,包括最早的脊索动物与各类奇异生物。
奥陶纪晚期,海洋生态系统已初具现代雏形。不久后,多细胞生物逐步征服陆地,生态空间进一步开拓。最终,陆地群落演化带动地球地球化学循环深刻转型,全球多样性与生产力由此大幅提升。
陆地生命的兴起
最早的陆地植物体型相对较小。大约3.8亿年前,最早的树木出现了。到3.5亿年前,由木贼、石松、蕨类、前裸子植物和种子植物组成的森林已经在全球广泛分布,覆盖了多个明显不同的生物群系。这对生物圈的影响似乎是巨大的。全球生产力飙升到前所未有的水平。煤炭以此后再也未达到的大量沉积。全球二氧化碳水平在大约5000万年内下降到之前的10%,最终稳定在接近现在的水平。这为随后时期的全球显著降温奠定了基础。
早期的陆地植物可能相对缺少天敌,比如植食性动物。到石炭纪中期,进攻已经开始了。具有特征性口器的昆虫、有咬痕的化石叶片,以及牙齿适合咀嚼的脊椎动物,都表明植物已经开始了与动物攻击的持续战争。这是一个重要的新营养级,因为现在植物生产力的一部分可以供其他生物利用了。陆地生态系统已经成熟。
飞行能力的演化
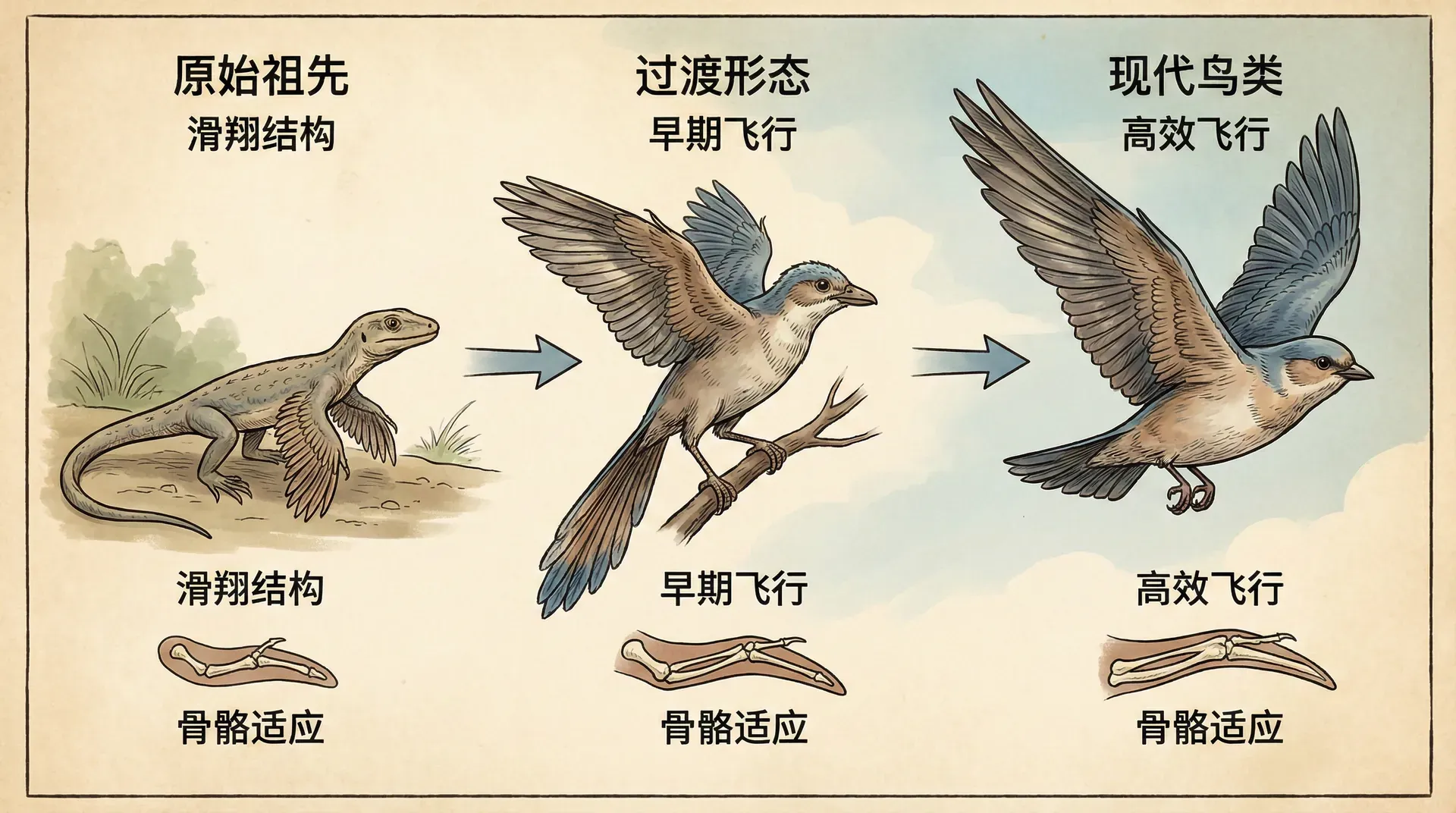
飞行被认为是生物演化史上最激动人心和深远的创新之一。在地球的历史长河中,“飞行”这一能力至少在四个动物类群中独立演化过,分别是昆虫、翼龙、鸟类和蝙蝠。这种独立演化的现象,被称为趋同演化。接下来,我们将以鸟类为核心,探究飞行能力是如何逐步演化出来的,并适度比较不同类群的演化路线与特点。
鸟类飞行的起源
现今多数古生物学家普遍认为鸟类起源于兽脚类恐龙,这是一类以双足行走、以肉食为主的恐龙。它们与现代鸟类之间,存在许多解剖特征上的联系。例如,兽脚类恐龙普遍具备相对较长并具灵活性的前肢、轻巧中空的骨骼、缩短和加固的尾巴、体型减小趋势以及可能拥有羽毛状皮肤结构。近几十年来,尤其是在中国辽宁所发现的多种带羽毛恐龙化石(如中华龙鸟、小盗龙),进一步证实了鸟类与恐龙之间的进化桥梁。
以中华龙鸟和小盗龙为代表的化石表明,羽毛最初可能用于身体保温或性状展示,而非专门为飞行而演化。
关于鸟类如何从地栖恐龙演变到能够飞行的动物,目前学界主要存在几种假说:
不同飞行动物的演化对比
无论是昆虫的薄翅,还是蝙蝠的飞行膜,不同类群都以极其不同的方式攻克了飞行的生物学难题。例如:
-
昆虫或许是最早掌握飞行能力的动物,其翅膀可能起源于祖先腿部靠近躯干的鳃状结构。从化石蜉蝣若虫到现代蜻蜓、金龟子,昆虫翅膀的多样结构成为动、静态飞行的典范。值得注意的是,现代昆虫有一套复杂的发育基因调控,只有胸部节段能够发育翅膀,但理论上身体其他节段也有潜力。
-
水生掠行起源假说:例如,有些石蝇和蜉蝣利用发育于胸部的“鳃翅”,在水面掠行并最终抵达陆地。这种结构最初用于呼吸,后经自然选择调整为飞行器官,兼具稳定和推进作用。
-
蝙蝠与翼龙的起源更为神秘。这两类脊椎动物至今尚未发现明确的过渡化石。它们共同的特征是:飞行膜由前肢及手指间的皮肤扩展形成,后肢灵活性较差,表明其飞行能力并非源自地面奔跑,而极有可能源于树栖滑翔的祖先。在蝙蝠的演化中,夜行和食虫也是典型特征,这有助于理解蝙蝠为何能遍布全球各地、生态类群极为丰富。
您可以通过下方的简单对比,看到四类飞行动物关键演化特征:
- 昆虫:翅膀由鳃状节段演化/身体小/外骨骼轻盈/早期出现在石炭纪。
- 翼龙:肢指间飞行膜/大脑容量增大/多为滑翔型飞行/三叠纪-白垩纪。
- 鸟类:羽毛前臂/坚固的龙骨突/高代谢/侏罗纪演化起源。
- 蝙蝠:手指支撑的飞膜/夜行/超声波回声定位/古近纪起源。
中国不仅拥有丰富的恐龙羽毛化石资源,也是蝙蝠多样性全球领先的国家之一,目前记录有超过130种本土蝙蝠,从热带雨林到城市都有分布。
演化时机的重要性
尽管现代生态中存在许多能够滑翔和短距离“飞行”的动物,例如飞鼠、飞蜥、飞鱼、树蛙等,但动力飞行(真正依靠挥动翅膀持续飞行)只有上述四类成功实现。那么,为什么会是这四类在特定时期形成飞行能力,而不是其他动物呢?背后的驱动力是什么?
从演化的角度来看,“重大功能创新”往往不是靠单一的突变实现,而是经由一系列微小、连续性的变异慢慢累积结果。例如,原始羽毛从简单的丝状结构到现代鸟类多类型的羽毛,演化上并无突兀的断裂,而是逐步排列组合。化石证据(如辽宁的带羽毛恐龙)很好展示了这一渐变过程。
外界环境因素同样不可忽视。近期学界普遍认为,大气氧气浓度的波动是推动飞行能力演化的关键。例如,晚石炭纪(约3.5亿年前)地球大气氧气含量曾达到巅峰,这一时期正是许多大型昆虫和第一批有翅昆虫化石出现的时间。之后的中生代(侏罗纪~白垩纪)氧气浓度再次上升,刚好与翼龙、鸟类的起源和多样化相吻合。到了古近纪(约6000万年前左右),蝙蝠也开始出现。
下表概括不同飞行动物出现的大致时间与相应氧气环境(并非所有时间节点都为峰值,仅作参考):
高氧气浓度对飞行生物有两个巨大优势:
- 空气密度增加,带来更高升力,使飞行更高效;
- 代谢水平提升,提供支持高能量消耗(如持续拍翼飞行)的能力。
此外,有学者提出,某些地质时期的环境变化(如由陆地化引起的生产力提升),间接推动了氧气浓度变化,而氧气浓度升高又反过来加速飞行能力的演化和创新。这种“演化—生态—环境正反馈”的模式可能在地球生物史中多次发生。
飞行能力的维持与生态影响
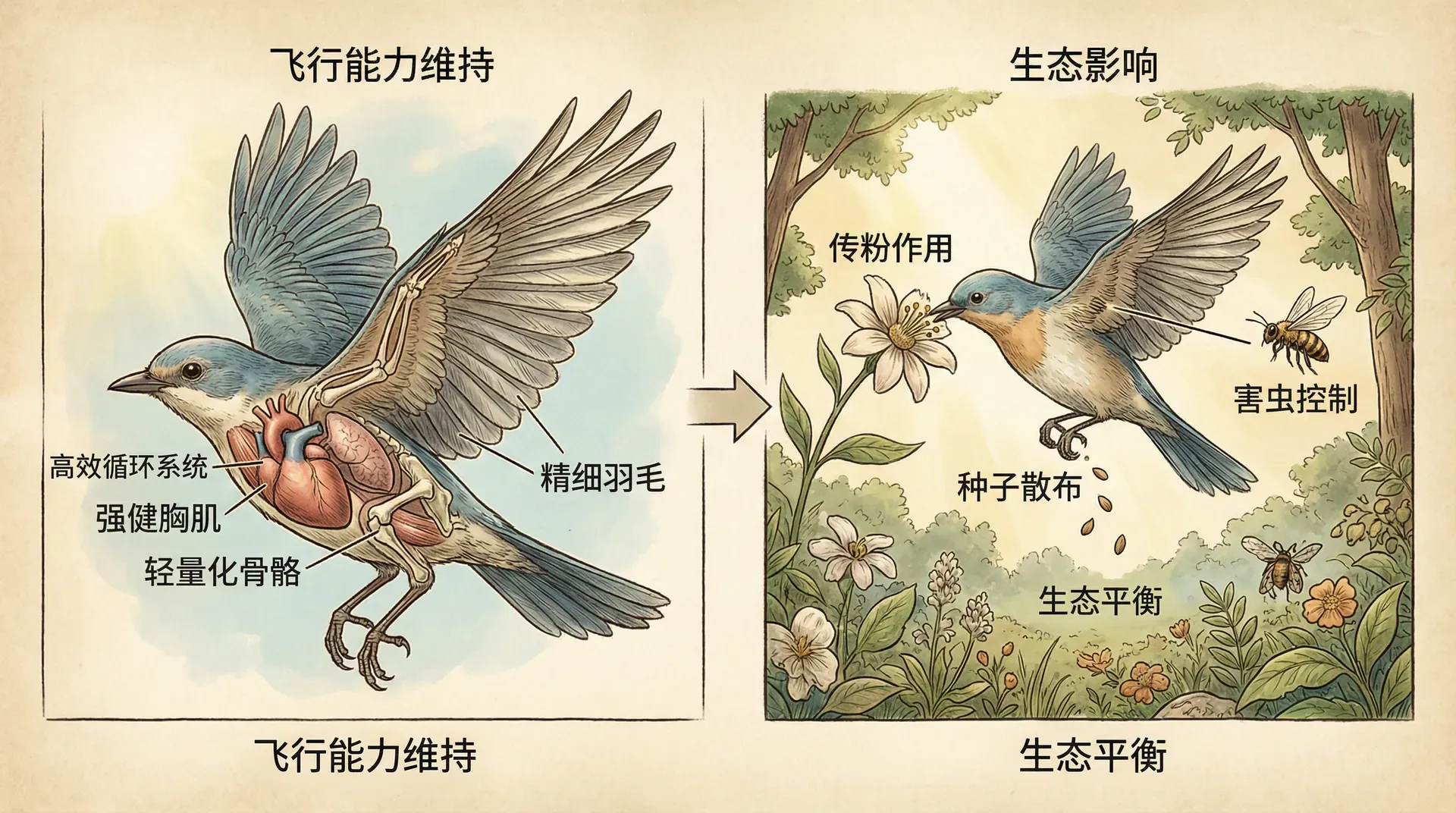
飞行起源后,只有长期维持下来,相关的生态影响才能持续体现。然而,飞行能力的丧失在不同类群中呈现不同频率:昆虫和鸟类中较为常见,翼龙和蝙蝠则几乎未见。其根本差异在于前者可分工翅膀与步行装置,后者则在地面上相对无效。
对于昆虫和鸟类来说,这使得在特殊环境下丧失飞行成为可能,尤其是在捕食压力较小的岛屿等地。例如,岛屿鸟类在缺乏天敌、局部高扩散死亡风险与食物断供等条件下,往往演化为不能飞。昆虫丧失飞行的情形更为普遍,甚至在除淡水以外的几乎所有生态系统反复发生。相比之下,寄生生活则更易导致昆虫失去飞行能力,典型代表如无翅的跳蚤和虱子。而总体来看,无翅分枝较少大辐射,究其原因或许与生态位转换和分化速率下降有关。
飞行与多样性和生态创新的关系
飞行不仅是生态空间的拓展(如“登陆”之后打开天空新世界),还影响了生物多样性和演化速度。主要体现在:
- 占据新生态空间:飞行让生物能利用空气层,开辟前所未有的生态位,极大推动多样化。
- 促进物种分化与分布:飞行提升迁徙与扩散能力,有助于隔离、分化及局部灭绝后的再定居。比如会飞的鸟类和蝙蝠普遍比同等体重哺乳动物寿命更长,这种生活史变化对灭绝率和物种形成都有潜在影响。
- 带动生态互作创新:飞行动物成为传粉等关键生态功能的主力,刺激了被子植物等类群的多样演化。例如中国云南的木兰科和甲虫、杜鹃与蜂蝶等即是明证。
在中国,许多特有植物类群与传粉者形成了精密的共生关系。比如云南的木兰科植物与甲虫的传粉关系,以及西南地区杜鹃花科与蜂类、蝶类的互动,都展示了飞行动物对植物多样性的深远影响。
飞行的演化贯穿了生态学重大转折的多个核心问题:从创新的起源、具体的演化时机与环境背景、关键功能的得失频率,到生态系统结构与物种多样性的变迁。我们看到,不同类群对飞行机制的依赖程度和丧失能力的频率差异极大,这影响了它们演化与生态命运。在未来,飞行这一主题或可与其他生态重大创新(如信息传递)一起,共同丰富我们对生态系统演化的理解和理论框架。