当生态遇上演化
在云南高原上,散布着一系列美丽的淡水湖泊。如果你在1960年代游览滇池,会惊讶地发现这片水域中生活着超过几十种土著鱼类,其中许多是滇池特有的物种,在地球上任何其他地方都找不到。这些鱼类包括金线鲃、滇池金线鲃、云南鲤等,它们各自占据不同的生态位,有的以浮游生物为食,有的啃食水底藻类,还有的捕食其他小鱼。
类似的故事也发生在青海湖。这个中国最大的内陆湖泊,海拔超过3000米,生活着一种特殊的鱼类——青海湖裸鲤。这种鱼在极端环境下演化出了独特的生理特征,能够在高寒缺氧的环境中生存繁衍。更有趣的是,在青海湖不同区域的裸鲤种群,已经开始展现出明显的形态差异。
特有种(endemic species)指的是只在某一特定地理区域自然分布的物种。云南高原湖泊因其地理隔离,孕育了大量特有鱼类。
让我们用数据来对比几个主要湖泊的鱼类多样性:
从上方表格可以看出,虽然青海湖面积最大,但物种多样性却相对较低,这与其高寒环境和相对年轻的地质历史有关。而云南几个高原湖泊虽然面积不大,却孕育了丰富的特有鱼类。
这些湖泊中的鱼类多样性是如何产生的?它们并不是从外部迁徙而来,而是在湖泊内部逐渐演化形成的。这个过程涉及生态学和演化生物学两个领域的深刻交互。
上图展示了滇池鱼类多样性的急剧下降。从1970年代开始,由于水质污染、外来物种入侵和过度捕捞,滇池土著鱼类经历了灾难性的衰退。这个真实的案例提醒我们:物种多样性的形成需要漫长的时间,但破坏却可能在几十年内完成。
物种是如何形成的
要理解湖泊中的鱼类多样性,我们首先需要搞清楚一个基本问题:新物种是怎么产生的?
地理隔离
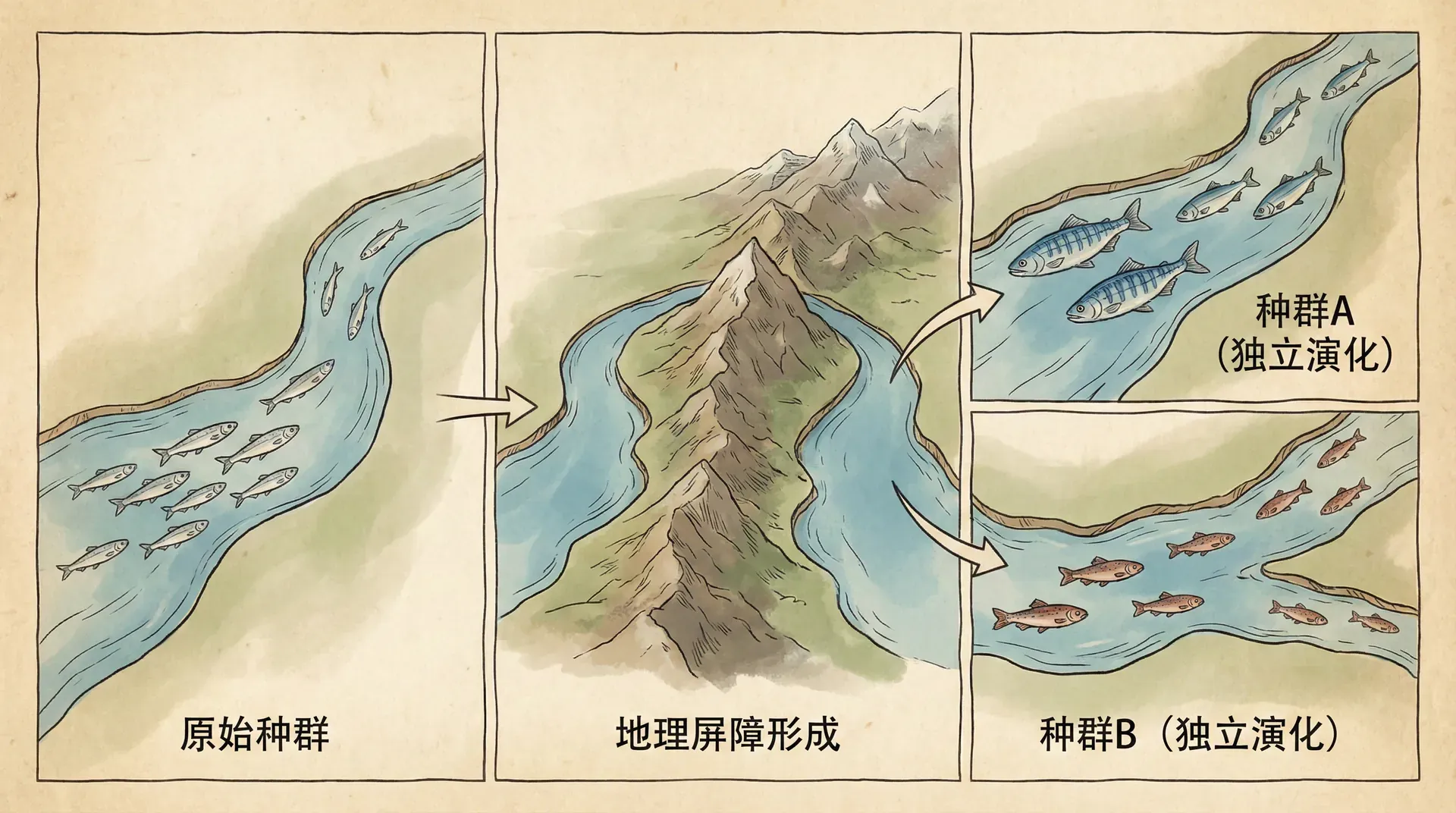
经典的物种形成理论认为,一个物种要分裂成两个或多个新物种,通常需要地理隔离。原本生活在同一片水域的鱼群,因为地质变化或者一小群个体被洪水冲到了新的湖泊,两个种群从此分离。在各自的环境中,它们面临不同的生存压力,经过成千上万代的自然选择,逐渐演化出不同的特征。最终,即使它们再次相遇,也已经无法互相繁殖了。
这个过程在地质历史上反复上演。青藏高原的隆升就是一个典型例子。随着高原抬升,许多水系被分割,原本连通的河流变成了相互隔离的水体。生活在不同水体中的鱼类开始各自演化。青海湖裸鲤的祖先本来是黄河水系的鱼类,大约在13万年前,青海湖与黄河水系分离,裸鲤种群被隔离在这个高原湖泊中,逐渐演化出了适应高寒环境的特征。
地理隔离导致的物种形成称为“异域物种形成”(allopatric speciation)。这是最常见也最容易理解的物种形成方式。
就地分化
但地理隔离并不能解释所有现象。在同一个湖泊中,为什么会存在这么多共同生活、却保持各自独立的物种?它们明明有机会相遇和交配,为什么没有混合成一个物种?
这就涉及到更有趣的机制了:在没有地理隔离的情况下形成新物种,我们称之为“同域物种形成”(sympatric speciation)。这个过程需要克服两个关键挑战:
-
生态分化:两个正在分化的种群必须占据不同的生态位,否则它们会因为竞争而无法共存。比如,一群鱼以浮游生物为食,另一群以藻类为食,它们虽然生活在同一片水域,但不会为同样的食物资源激烈竞争。
-
生殖隔离:即使两个种群占据不同的生态位,如果它们还能自由交配,基因就会不断混合,无法形成独立的物种。所以必须有某种机制阻止或减少种群间的交配。
同域物种形成的关键在于,既要有生态上的差异化,又要有生殖上的隔离机制。这两个过程必须协同发生。
配偶选择的力量
在鱼类中,一个重要的生殖隔离机制是配偶选择。许多鱼类在繁殖季节会展现出鲜艳的婚姻色,雄鱼用这些色彩吸引雌鱼。如果雌鱼有强烈的颜色偏好,比如有些雌鱼特别喜欢红色雄鱼,而另一些偏爱蓝色雄鱼,那么久而久之,种群就可能分裂成不同的类型。
假设一个湖泊中的鱼类原本颜色都比较普通,是灰褐色的。某些雄鱼突变产生了轻微的红色,另一些产生了轻微的蓝色。如果雌鱼的视觉系统对红色和蓝色特别敏感(这可能是由于视觉色素的遗传特性),那么她们会对这些有颜色的雄鱼表现出偏好。
说明:最初的鱼群为中性色,部分个体突变出蓝色或红色,雌鱼根据视觉偏好与同色雄鱼交配,逐步形成两个独立物种。
偏好红色的雌鱼主要与红色雄鱼交配,它们的后代既继承了红色基因,也继承了偏好红色的基因。同样,偏好蓝色的雌鱼主要与蓝色雄鱼交配。随着时间推移,这两个群体的基因交流越来越少,最终形成两个独立的物种。这个过程被称为“失控选择”(runaway selection),因为一旦开始,就会自我强化。
一旦通过配偶选择形成了生殖隔离,两个种群就可以各自适应不同的食物资源或栖息地,发生生态分化。这样,同域物种形成就完成了。
环境因素的关键作用
物种的形成和维持不仅取决于生物本身的遗传和行为特性,还受到外部环境条件的深刻影响。环境不仅提供了生存空间和资源,还经常决定了哪些物种能够出现、多样化并长期稳定地共存。水体的温度、透明度、营养情况、光照等一系列生态因子,都可能深刻影响生物多样性的形成、维持和演化。因此,环境条件在物种形成过程中常常起到决定性甚至主导性的作用。
水质透明度的影响
在研究湖泊鱼类时,科学家发现了一个有趣的规律:在水质清澈的区域,鱼类物种多样性往往更高,而且鱼的颜色更加鲜艳;在浑浊的水域,物种数量减少,鱼的颜色也变得暗淡。
为什么会这样?原因就在于视觉通讯。如果水质浑浊,雌鱼很难看清雄鱼的颜色,基于颜色的配偶选择就会失效。当选择压力消失,不同物种之间的生殖隔离就会崩溃,它们开始混合交配,原本独立的物种又融合成一个。
以上信息说明了水体透明度与鱼类物种多样性的关系。透明度用塞氏盘深度表示——将一个黑白相间的圆盘沉入水中,还能看见的深度就是透明度(单位:厘米)。可以看到,随着水质变清澈,共存的物种数量显著增加。
这给我们一个深刻的启示:环境质量不仅影响物种能否生存,还影响物种能否保持各自的独立性。滇池的悲剧不仅是鱼类数量的减少,更是物种多样性的消失。当水体富营养化导致水质浑浊时,不同物种之间的界限变得模糊,特有种群消失了。
环境退化对生物多样性的影响是多方面的,它不仅直接导致物种死亡,还会破坏维持物种多样性的机制,让物种之间的界限消失。
生态机会的作用
为什么有些生物类群特别容易产生多样性,而另一些则相对单调?除了配偶选择,生态机会也很重要。
许多鲤科鱼类具有特殊的咽喉齿结构——这是位于喉部的第二套“颚”,专门用来研磨食物。这种结构让口部可以专门负责捕获食物,而咽喉齿负责加工食物,两个系统各司其职。这种“解耦”的设计大大增加了食性的可塑性。同一种鱼的不同种群可以快速适应不同的食物类型,从吃软体藻类到啃硬壳螺蛳,从过滤浮游生物到捕食小鱼,都能胜任。
这种生态灵活性意味着,当鱼类进入一个新的湖泊环境时,可以迅速分化去利用各种可用资源。如果同时再加上强烈的配偶选择,物种形成的速度就会非常快。
生态与演化的交响
现在我们可以回到最初的问题:为什么要把生态学和演化生物学放在一起讨论?
两个领域的交汇
生态学研究生物与环境的相互作用。它关心的问题包括:为什么某种生物生活在这里而不是那里?一个地区能容纳多少个体?哪些物种会共同生活?生态系统是如何运转的?
演化生物学研究生物谱系如何随时间变化、分支和灭绝。它关心的问题包括:某种特征是如何演化出来的?为什么某些类群特别多样?物种是如何起源的?
这两个领域看似关注不同的问题,但它们紧密相连,因为:
生态与演化的关系是双向的:生态过程是演化的动力,演化结果决定生态格局。理解一个领域能够帮助理解另一个领域。
从个体到生态系统
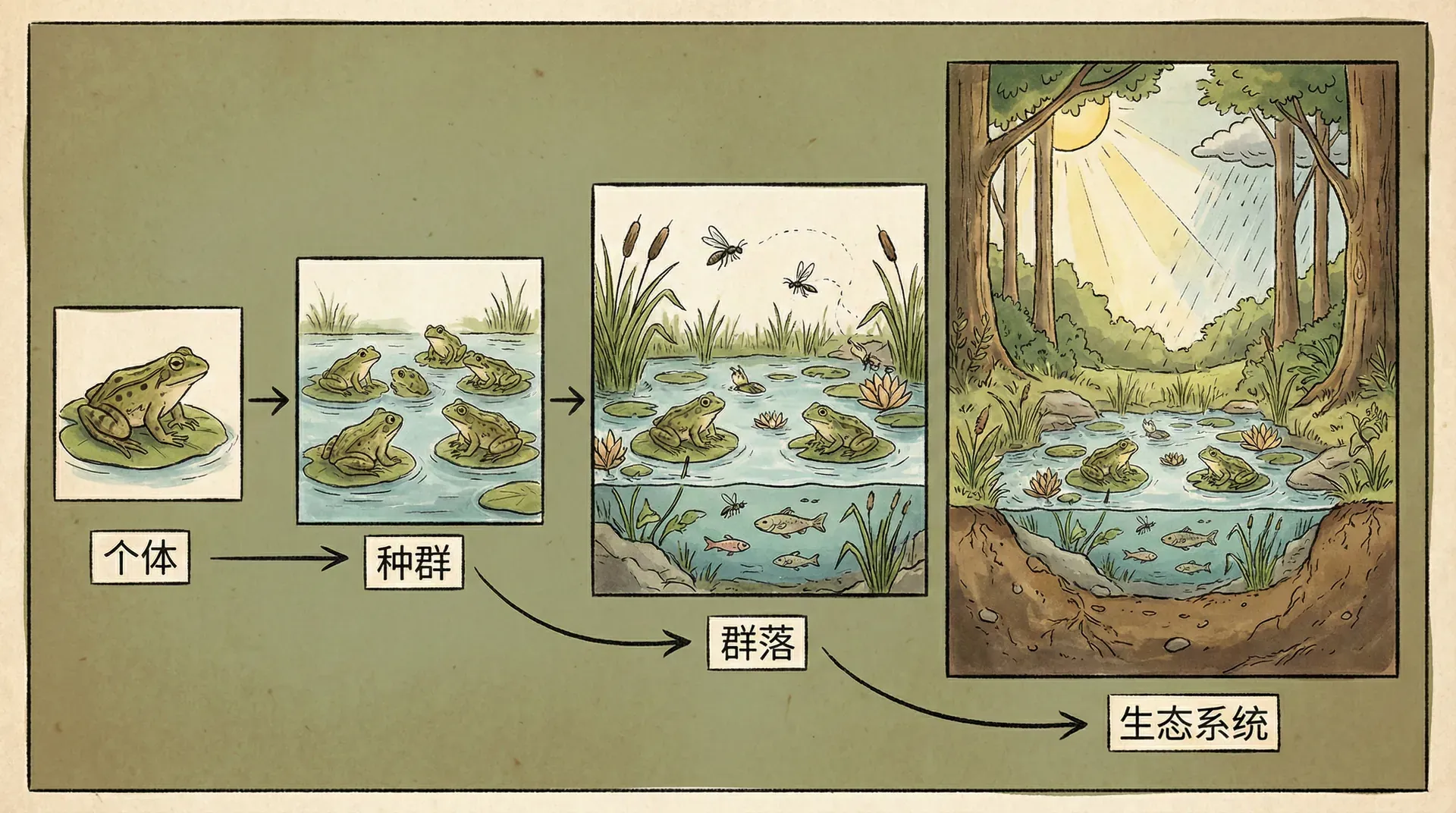
让我们看看演化如何影响不同层次的生态学问题。
个体层面:为什么某种鱼长着特定形状的颚?为什么在繁殖季节展现鲜艳的色彩?为什么偏好某种配偶?这些都是演化的结果。行为生态学专门研究生物的行为如何适应环境——什么样的行为策略在什么样的环境中最有利。我们讨论的配偶选择行为,就是行为生态学的经典问题。
种群层面:一个物种能有多少个体,种群数量如何波动,这取决于该物种的生活史特征——多久繁殖一次、一次产多少卵、寿命多长。这些特征都是长期演化的结果。有些鱼类产卵数量少但亲鱼会照料幼鱼,另一些鱼类产卵数量巨大但完全不管后代,这两种策略各有利弊,适应不同的生态环境。
群落层面:一个湖泊能容纳多少物种,这些物种如何相互作用?这既取决于生态因素(有多少种资源、环境有多稳定),也取决于演化因素(物种形成的速度、生态特化的程度)。我们看到,具有特殊性状(如解耦的咽喉齿、强烈的配偶选择)的类群更容易产生高度多样的群落。
生态系统层面:物种的功能特征决定了生态系统如何运转。食物网的结构、营养物质的循环、能量的流动,都与物种的演化特征密切相关。
从湖泊到世界
云南湖泊的鱼类只是一个小小的例子,但它展示的原理具有普遍性。在世界各地,我们都能看到生态与演化的交互作用。东非大湖泊的慈鲷鱼,加拉帕戈斯群岛的达尔文雀,夏威夷群岛的果蝇,每一个案例都讲述着相似的故事:在特定的地理和生态条件下,在特定的演化机制作用下,生命迸发出惊人的多样性。
理解这些过程不仅满足我们的好奇心,也有实际意义。当我们知道物种多样性是如何产生和维持的,就更能理解如何保护它。滇池的教训告诉我们,破坏生态环境不仅会导致个别物种灭绝,还会摧毁整个演化过程。我们失去的不仅是现在的物种,更是未来产生新物种的可能。
保护生物多样性,不仅要保护现有物种,还要保护产生和维持多样性的生态与演化过程。
总结
物种形成常常通过地理隔离或者在同一环境中的分化来实现。例如,青海湖的裸鲤因为隔离而产生演化分化,而同一湖内的不同鱼类则可能通过配偶选择逐步分化成新的物种。配偶选择在物种分化中也扮演着重要角色,比如雌性偏好特定颜色的雄性会导致群体内出现生殖隔离,从而推动新物种的产生和进化。
此外,环境条件对物种的形成和维持起着决定性影响。以水质清澈度为例,它直接影响鱼类的视觉通讯方式,进而影响物种的多样性。某些特殊性状也会为物种提供更多的生态机会。例如,鲤科鱼类拥有“解耦”的咽喉齿,这一特点让它们能够适应多样化的食物资源,增强对生态环境的适应能力。
最后,生态过程与演化过程始终是相互作用、相互推动的。生态因素往往会通过自然选择改变生物性状,而这些性状的变化又会进一步决定生物在生态系统中的角色和地位。通过理解这些核心概念之间的关系,我们能够更好地把握生物多样性的本质与维持机制,揭示生命世界的复杂性。