适应性生存策略

当我们走在田野里,看到向日葵随着太阳转动,或者观察爬山虎沿着墙壁攀爬,可能会觉得这只是植物的本能反应。但实际上,植物拥有一套精密的生存策略,它们能够感知环境变化,并做出相应的调整。这些调整有时甚至比我们想象的更“聪明”。
虽然植物没有神经系统,不能像动物那样快速移动或做出复杂的行为,但它们通过改变生长方向、调整资源分配、甚至选择性地放弃某些后代,来最大化自己的生存和繁殖机会。这些策略在漫长的进化过程中被不断优化,形成了今天我们看到的各种奇妙现象。
接下来,我们将从三个角度来理解植物的适应性策略:如何获取资源、如何处理家族关系、以及如何在繁殖中竞争。这些看似只适用于动物的概念,在植物世界中同样精彩。
资源获取的智慧
投资决策
当你在超市购物,手里的购物车已经装了一些商品。你是继续在这个货架前挑选,还是去下一个货架?这个决策取决于两个因素:这个货架还有多少你想要的东西,以及走到下一个货架需要多长时间。如果这个货架的好东西已经被你挑得差不多了,那么即使走到下一个货架需要花点时间,你也应该离开了。
这个简单的道理,在生物学中有个专门的名字,叫做“边际价值定理”。对于动物来说,它们在不同的食物区域之间移动,需要决定在每个区域停留多久。对于植物来说,虽然它们不能移动整个身体,但它们的根系、藤蔓或枝条可以生长到不同的区域,同样面临资源分配的问题。
菟丝子的选择
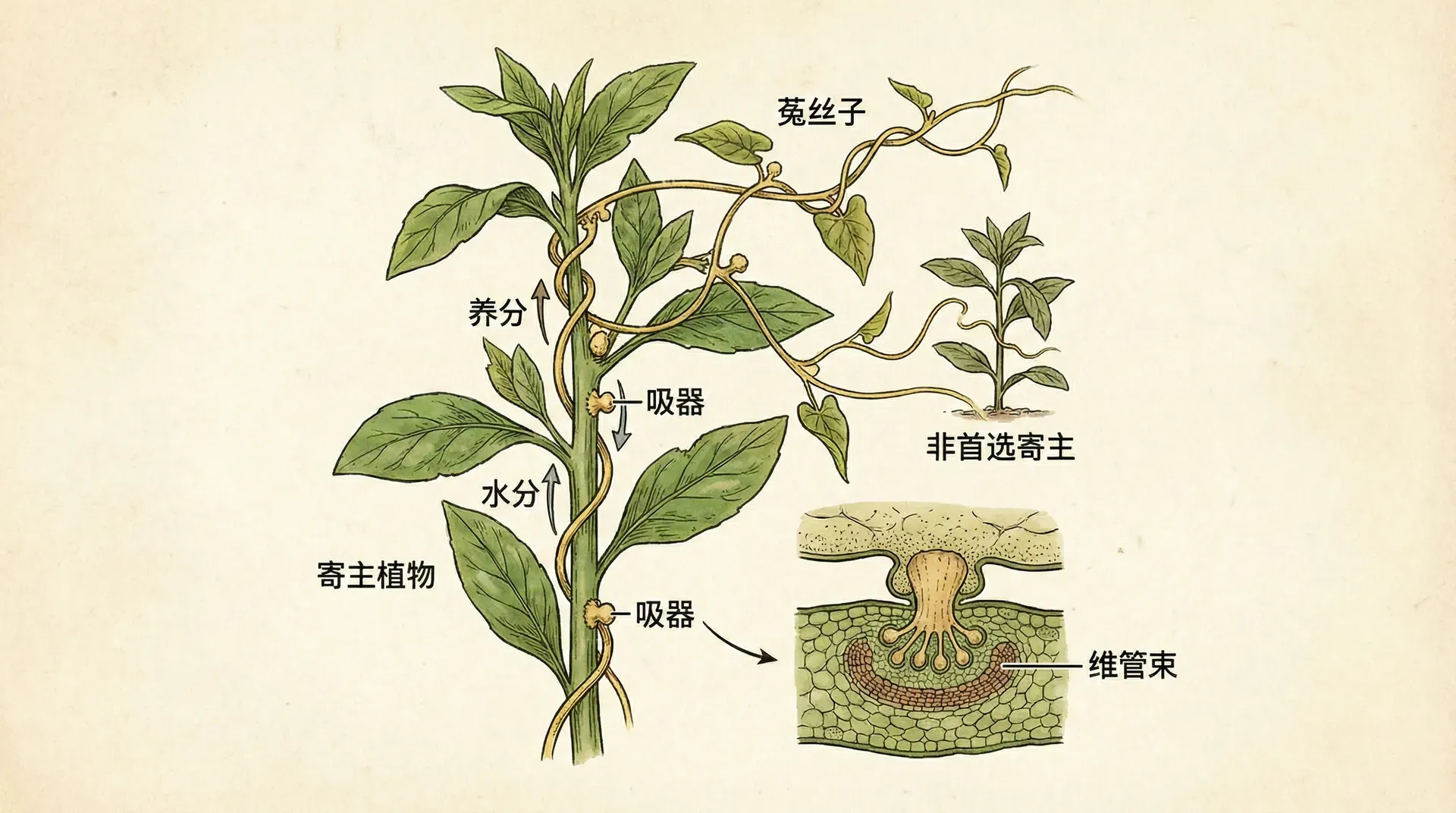
中国的田野和山区常见一种奇特的植物——菟丝子。它没有根,也没有叶片,更不能进行光合作用,完全依靠缠绕在其他植物茎干上,用特殊的吸盘吸取宿主的养分和水分。菟丝子的茎干是金黄色的细线,远远看去像给宿主植物披上了一层金色的网。
菟丝子一生中会遇到许多不同的宿主植物,有的营养丰富(比如豆科植物),有的营养贫乏(比如某些杂草)。它该如何选择?研究发现,菟丝子会在营养好的宿主上缠绕更多圈,形成更密集的吸盘;而在营养差的宿主上,它只是简单地搭个桥,然后继续生长寻找下一个宿主。
这完美符合边际价值定理的预测。对菟丝子来说,“旅行时间”就是从一个宿主生长到下一个宿主所需的时间和能量,“停留时间”就是在某个宿主上缠绕的圈数。在宿主密度高的环境中,菟丝子可以快速找到下一个宿主,因此它在每个宿主上的投资比较少;而在宿主稀疏的环境中,菟丝子会在每个遇到的宿主上缠绕更多,因为找到下一个宿主的成本太高了。
农作物根系的策略
如果说菟丝子的例子还有些特殊,那么我们来看看最普通的农作物——小麦和水稻。它们的根系面临一个类似的问题:土壤中的养分分布并不均匀,有的区域肥沃,有的区域贫瘠,根系该如何分配呢?
科学家通过实验发现,当土壤被人为分成不同养分浓度的区域时,植物根系会优先在养分丰富的区域生长。而且,土壤养分差异越大,根系分布的差异也越大。这说明植物能够感知土壤养分的差异,并相应地调整根系的生长方向和密度。
假设有两个土壤区域,一个养分浓度高,一个养分浓度低。下图显示了根系投资(根的密度或重量)与养分吸收率的关系:
可以看到,在高养分区域,每单位根系投资带来的回报更高,因此植物会将更多的根系分配到这些区域。但同时要注意,即使在高养分区域,收益曲线也会逐渐趋于平缓,这意味着过度投资也不划算。植物需要在不同区域之间找到一个平衡点,使得总的养分吸收率最高。
不同植物的资源获取策略
不同植物采用的策略会有所差异,这取决于它们的生长习性和所处的环境。下表总结了几种常见植物的资源获取特点:
边际价值定理告诉我们:生物在资源获取时,不是简单地“有资源就利用”,而是要计算投入产出比。当某个资源区的边际收益降到平均水平以下时,就该转移到下一个区域了。这个原理不仅适用于动物觅食,也适用于植物的生长分配。
家族中的合作与冲突
亲缘关系的分量
在人类社会中,我们对家人往往比对陌生人更好。这背后有深刻的生物学原因。20世纪60年代,生物学家汉密尔顿提出了一个简洁而深刻的规则,用来预测什么时候利他行为会被自然选择青睐。这个规则用数学表达就是:
这里的 是受益者获得的好处, 是行为者付出的代价,而 是亲缘关系系数。亲缘关系系数衡量的是两个个体共享基因的概率。对于有性繁殖的生物来说,父母和子女之间的 (因为子女有一半基因来自父母),同父同母的兄弟姐妹之间也是 ,而祖孙之间是 。
这个不等式的含义是:只有当受益者得到的好处乘以亲缘关系系数,超过行为者付出的代价时,这种利他行为才会进化出来。换句话说,关系越近,越容易出现利他行为;关系越远,就需要受益者得到更大的好处才行。
但这个规则还有另一面:它也定义了自私的界限。即使是亲兄弟姐妹,如果资源的价值对双方一样(),那么每个个体还是会优先把资源留给自己,而不是兄弟姐妹。因为 ,不等式 不成立。这就是为什么即使在亲密的家庭中,也会存在竞争和冲突。
下方图表理解不同亲缘关系的 值:
植物的家族问题
植物也有丰富的家族生活吗?当然有。一株开花植物的果实里,往往包含多个种子,这些种子就是兄弟姐妹。它们共享同一个母亲(提供果实和养分的植株),但可能有不同的父亲(不同来源的花粉)。在一个果实的狭小空间里,这些种子要竞争有限的养分,这就是典型的家族冲突场景。
更有趣的是,植物的种子并不是简单的胚胎,而是一个复杂的结构。以玉米为例,一粒玉米由三个主要部分组成:胚(将来发育成新植株)、胚乳(储存养分供胚发育)、种皮(保护结构)。其中,胚是二倍体,包含父母各一半的基因;而胚乳是三倍体,包含母本两份基因和父本一份基因(2母:1父)。
这个看似奇怪的安排,实际上是家族冲突的产物。我们来算算不同成员之间的亲缘关系:
胚乳的三倍体结构(2母1父)使它的利益更接近母本,而不是单纯的胚。胚乳中母本基因占2/3,这让母本能够通过胚乳更好地控制养分分配,避免某个胚过度自私。这是进化过程中母本对父本的一种"制衡"机制。
水稻的选择性流产
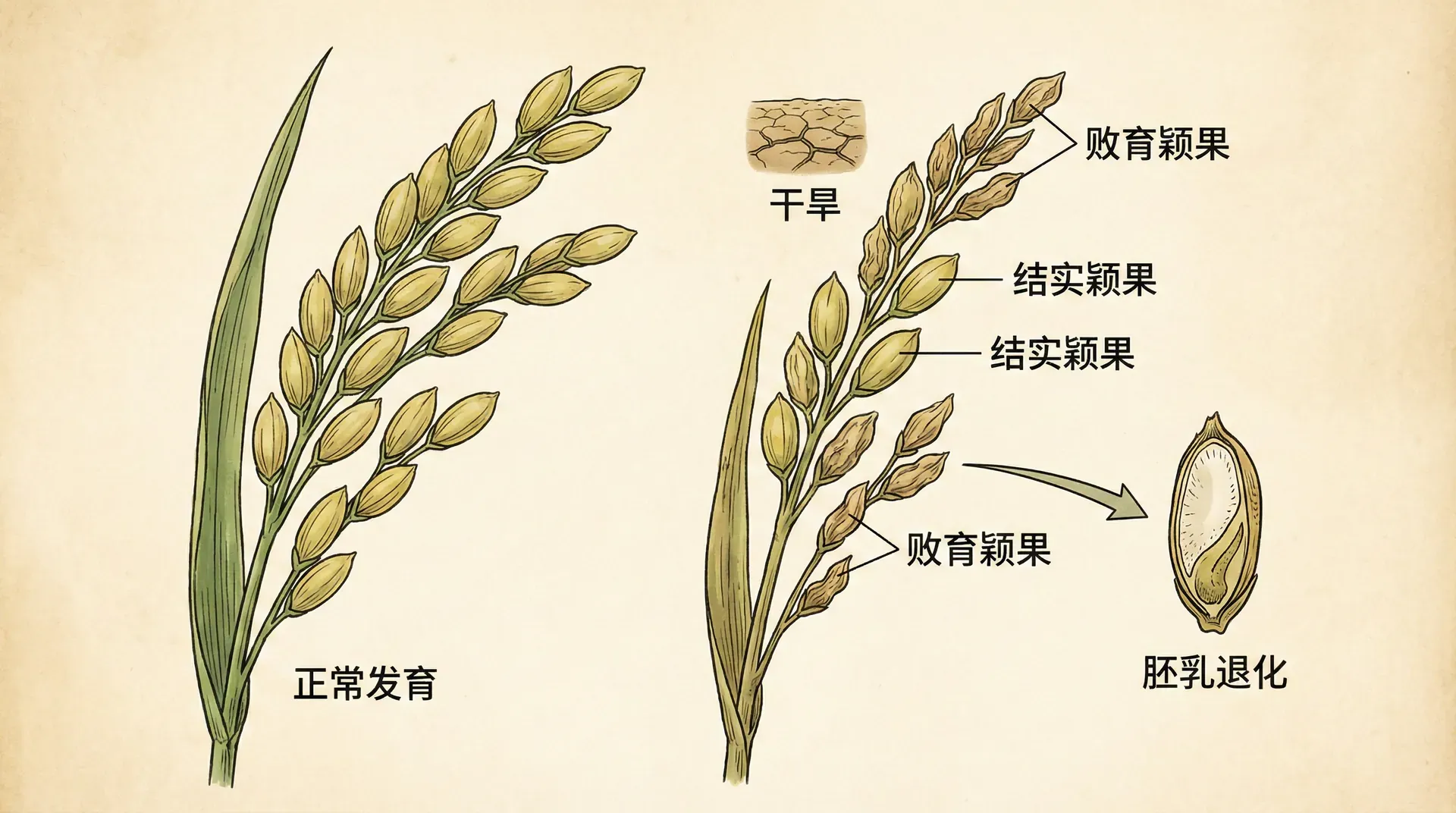
在湖南的水稻田里,农民们发现一个有趣的现象:同一株水稻的谷粒成熟度并不一致,有些饱满充实,有些却干瘪空瘪。仔细观察会发现,那些空瘪的谷粒往往位置不佳(比如离主茎较远),或者受粉时间较晚。
这不是病害,而是水稻主动的选择。水稻母本会评估每个胚的“质量”,包括它的遗传背景、发育时机和位置。那些质量不佳的胚会被选择性地中止供应养分,导致种子发育不全或流产。这对母本是有利的,因为她的资源有限,与其平均分配给所有后代(包括质量差的),不如集中投资给那些更有希望的后代。
研究发现,水稻能够识别并选择性地流产自交的种子。自交种子的遗传多样性低,更容易积累有害突变,因此竞争力较弱。母本通过流产这些种子,提高了整体后代的质量。在实验中,如果人工授粉让水稻进行自交,流产率会明显上升;而异交授粉的种子则有更高的保留率。
类似的现象在玉米中也有发现。在四川的玉米田里,如果土壤中铜含量过高(可能来自矿区污染),玉米能够选择性地流产那些对铜敏感的种子,保留那些耐铜的种子。这意味着母本不仅能够识别种子的杂合度,甚至能够识别某些特定的生理性状。这种能力帮助植物在恶劣环境中提高后代的存活率。
桃树的疏果智慧
在江南的桃园里,每年春天桃花盛开后,会结出密密麻麻的小桃子。但到了初夏,果农会发现地上落了一层小青桃,这是桃树自己掉落的。即使不算这些自然落果,果农还要人工疏果,把一部分小桃子摘掉,只留下间距适当的果实。
为什么要疏果?因为桃树的养分供应是有限的。如果让所有的果实都继续生长,最后会得到一树小而酸的桃子,没有商品价值。通过疏果,留下的果实能够获得充足的养分,长成个大、甜度高的优质桃子。
但在果农人工疏果之前,桃树自己已经先疏了一轮。这说明桃树母本也知道这个道理:与其养一堆质量差的后代,不如集中资源养少数优质后代。自然落果往往是那些授粉不良、位置不佳、或者遗传质量有问题的果实。母本通过某种机制识别出这些“问题儿童”,主动中断对它们的营养供应。
有趣的是,如果果实之间距离太近,落果率会更高。这可能是因为果实之间会产生竞争信号,甚至可能释放化学物质相互抑制。从进化角度看,这种兄弟相残的行为虽然残酷,但确实有利于提高整体的繁殖成功率。因为两个太近的果实会互相遮挡阳光,而且成熟后也会互相挤压,降低种子的传播效率。
母本的控制权
从上面的例子可以看出,在植物的家族冲突中,母本往往掌握着最终的控制权。这是因为种子发育所需的养分都要经过母本的输导组织,母本可以选择给谁、给多少。种皮(包裹种子的外壳)也是母本组织,这进一步加强了母本的控制。
但这不意味着父本和后代就毫无发言权。胚乳的三倍体结构就是一种妥协。某些植物中还发现了“基因组印记”现象:同一个基因,如果来自父本,表达水平就高;如果来自母本,表达水平就低(或者相反)。这是父母双方在基因水平上的博弈,就像在谈判桌上争夺话语权。
植物的繁殖竞争
花朵的展示
每年三月,江西婺源的油菜花开了,金黄色的花海吸引着无数游客和蜜蜂。四月,山东泰安的桃花盛开,粉色的花朵开满整个山谷。这些美丽的景象背后,是植物在繁殖中的激烈竞争。
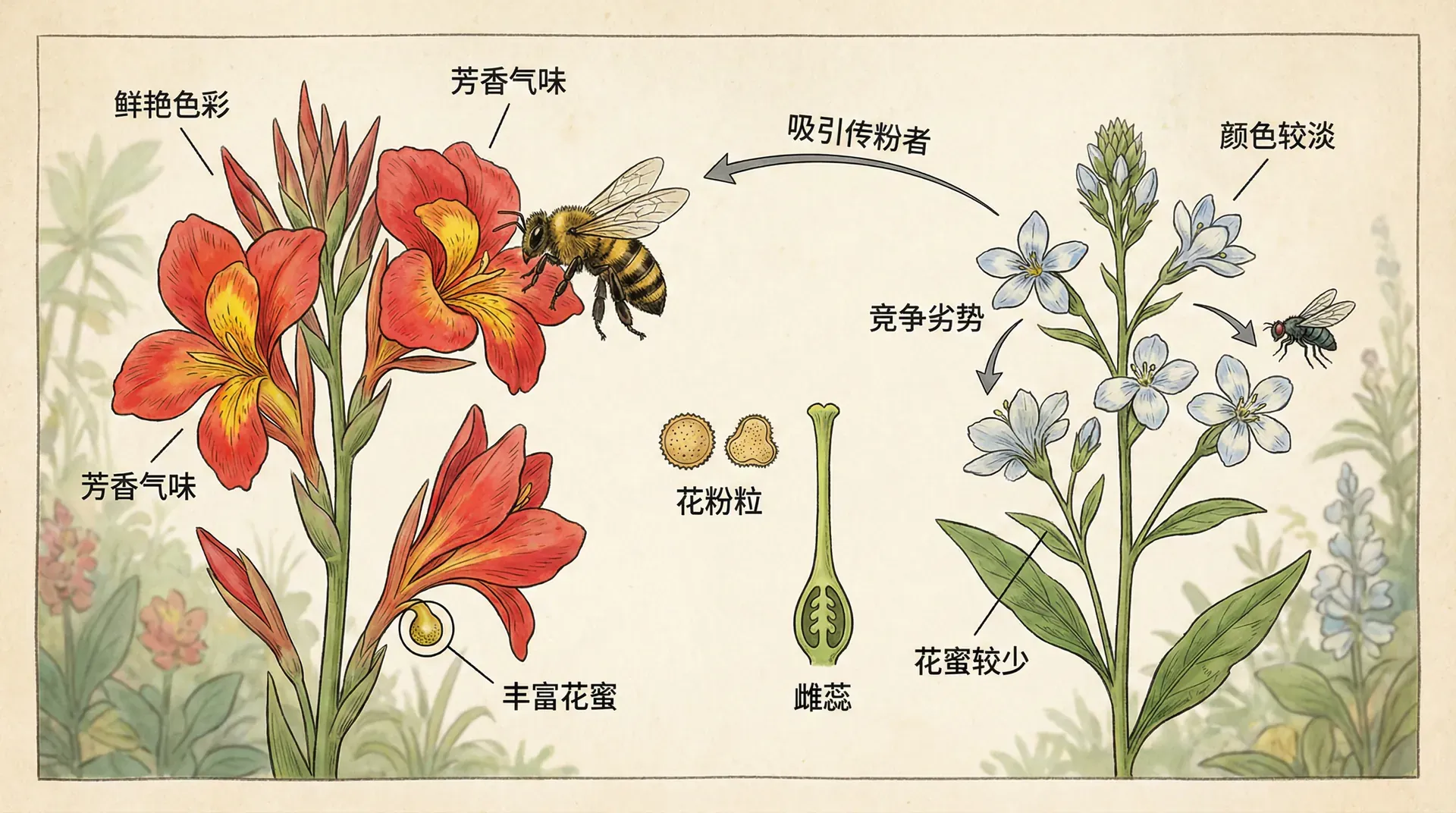
对于依赖昆虫传粉的植物来说,能否吸引到传粉者,直接决定了繁殖成功率。花朵就是植物的“广告牌”,颜色越鲜艳、花朵越大,就越容易吸引传粉者的注意。但是,制造和维持大而艳丽的花朵需要消耗大量的能量和资源。因此,花朵大小和数量之间存在权衡。
有些植物采取的策略是:一开始开很多花,远远看去非常壮观,吸引大量传粉者前来。等传粉者到了,再逐步减少开花数量,甚至让一部分花凋落。这些"牺牲"的花朵实际上充当了“诱饵”,它们的使命不是结果,而是吸引传粉者。这种现象在学术上叫做“雄花功能”,因为这些花虽然有雌蕊,但实际上主要功能是帮助散发花粉。
在西南地区常见的龙舌兰科植物中,这个现象特别明显。一个花序上可能有上百朵花,但最后只有十几朵能结果。那些不结果的花并不是失败者,而是为了整株植物的繁殖成功而做出的战术性牺牲。它们通过提供花蜜吸引蜜蜂和蝴蝶,从而增加整个花序的花粉传播量。
花朵数量与繁殖成功的关系
下图展示了不同植物的花朵投资策略:
从表中可以看出,不同传粉方式的植物,其花朵投资策略差异很大。虫媒植物需要花大力气吸引昆虫,而风媒植物则靠数量取胜。自花授粉植物因为不需要传粉者,所以花朵最为节俭。
花粉竞争
如果说花朵的展示是公开的竞争,那么花粉的竞争则是隐蔽的战场。当多个花粉粒落在同一个柱头上时,它们会展开激烈的竞争:谁能最先长出花粉管,到达子房,完成受精,谁就是赢家。
花粉管的生长是一个需要资源和能量的过程。有趣的是,提供这些资源的,恰恰是雌性的花柱组织。也就是说,雌性不仅是被动的接受者,她还控制着竞争的场地。在某些植物中,花柱会在受粉后变窄,这增加了花粉管之间的竞争强度,因为空间和资源都变得更加有限。
研究发现,桃树的花柱在受粉后基部会收缩,这减少了能够同时生长的花粉管数量,迫使它们更加激烈地竞争。这种机制帮助雌性筛选出生长最快、质量最好的花粉,从而提高后代的质量。
更神奇的是,花粉的竞争力有时候能反映父本的整体质量。在一些紫罗兰的研究中发现,那些产生高质量种子的父本植株,其花粉管生长速度也更快。这意味着,通过选择生长快的花粉,雌性间接地选择了基因质量更好的父本。这类似于动物世界中的“好基因”假说:雌性通过雄性的某种性状(如孔雀的尾巴),来判断其整体的基因质量。
植物的性选择虽然不像动物那样显眼,但同样精妙。花朵的展示吸引传粉者,这是“雄性竞争”;花粉管的竞争和选择性流产,这是“雌性选择”。两种机制共同作用,提高了植物的繁殖成功率和后代质量。
总结
植物在自然选择下演化出多样而高效的生存策略。它们不仅主动分配资源,在根、茎、叶等器官间灵活调整,比如菟丝子能辨别宿主、农作物根系因土壤养分而变化,实现收益最大化;还通过选择性流产或差异供给保障优质后代的生长,家族成员间也会为资源展开竞争与协调。
在繁殖方面,植物进化出各种吸引传粉者的机制,如色彩鲜艳、香气四溢的花朵,并通过花粉间的激烈竞争和花柱的筛选机制,择优产生后代。无论是公开的“展示”还是隐秘的“竞争”,都体现了其提升繁殖效率和后代质量的精妙设计。
这些看似简单的生长行为,其实源自长期进化积累的智慧:植物无需大脑,通过基因与自我调控,精准适应并优化环境,是“无声的智慧生命”。理解这些策略,不仅有助于生态保护和农业提升,也让我们看到生命各层面的共同法则——适应、竞争与优化。下次走进自然时,请用心观察那些宁静的植物——它们正上演着复杂而精彩的生存博弈,值得我们敬畏与深思。