演化与种群数量
在春季的鄱阳湖畔,成千上万只候鸟陆续飞来。你会注意到,有些鸟选择在水草丰美的区域落脚,有些则去了看起来条件一般的滩涂。这些鸟是随便找个地方停下的吗?它们的分布模式会不会影响整个种群的存活?当环境条件改变时,生物会怎样应对?而它们的演化又会如何影响种群的命运?
演化与生态之间存在双向作用:环境选择塑造生物特征,而生物特征的改变又会影响种群动态,进而改变环境条件,形成反馈循环。
生物对环境的适应会以两种方式影响种群数量:
-
生物表现出灵活的行为策略,这些策略其实是过去演化积累的结果。候鸟选择在哪片湿地越冬,这种决策能力经历了漫长的演化过程。虽然我们观察的这段时间里看不到明显的进一步演化,但这种行为本身就在影响种群如何分布。
-
演化可能就发生在生态学关心的时间尺度上,也就是几十年、甚至更短的时段内。这样的话,我们就得同时追踪演化和生态两个过程的动态了。
动物如何选择栖息地
让我们从一个具体问题开始:当一大群动物面对质量不同的栖息地时,它们会如何分布?
理想自由分布的逻辑
假设现在有一片湿地系统,包含了几块质量不同的区域。第一只到达的候鸟会怎么做?很自然地,它会选择食物最丰富的那块区域。但是,当第二只鸟到来时,情况就变了。第一只鸟已经在最好的区域觅食,消耗了一部分食物,也可能干扰到后来者的觅食。所以对第二只鸟来说,最好的区域现在看起来没那么好了。
随着越来越多的鸟抵达,最好的区域逐渐被占据,直到它的实际觅食收益降到和第二好的区域一样。这时候,新来的鸟就会在两个区域之间交替选择。最终,如果所有的鸟都能准确评估各区域的食物量和竞争者数量,并且可以自由移动,那么它们会达到一个稳定的分布状态——没有任何一只鸟能通过换到别的区域而获得更高的收益。
这种分布被称为“理想自由分布”(Ideal Free Distribution),是一种演化稳定策略。在这个状态下,每个栖息地的个体数量和它的生产力成正比,最好的地方聚集最多的个体。
从图中可以看到,最初优质栖息地能提供更多食物(绿线起点较高),但随着鸟类数量增加,竞争加剧,每只鸟的收益下降。当两条曲线相交时,两个栖息地的收益相等,这就是平衡点。此后,鸟类会在两个区域间分配,保持收益相等。
预测栖息地变化
这个理论不只是纸上谈兵。让我们看一个实际问题:如果湿地面积减少了,候鸟种群会发生什么变化?
以鄱阳湖的越冬候鸟为例。每年秋天,大量的鸿雁、豆雁从西伯利亚飞到这里过冬。假设因为人类活动,部分湿地被占用,可供觅食的区域缩小了10%。我们能预测种群数量会下降多少吗?
预测的逻辑是这样的:当湿地面积减少,同样数量的鸟被挤进更小的空间,密度增加。根据理想自由分布模型,我们可以计算出在新的、更小的栖息地中,鸟类密度和食物获取率的关系。食物获取率降低意味着越冬死亡率上升。另一方面,我们也需要考虑繁殖地的情况,建立出生率与密度的关系。当出生率恰好平衡死亡率时,种群就达到平衡。
有研究表明,基于行为模型的预测往往比单纯的人口统计模型更准确。原因有两个:
-
观察个体行为相对容易,而直接测量种群层面的出生率和死亡率既费时又费力。
-
更重要的是,当环境改变时,密度-死亡率的直接关系也会改变,但动物的行为策略本身相对稳定。
通过研究行为机制,我们实际上是在研究变化背后的根本原因,因此能更好地预测新环境下会发生什么。
实际应用中,这类“基于行为的模型”已经用于预测人类干扰、栖息地丧失、海平面上升等对鸟类和哺乳动物种群的影响。关键在于,我们假设这些适应性行为是过去演化留下的遗产,生物已经拥有应对环境变化的能力。但还有另一种可能:生物正在我们眼前演化。
快速演化就在眼前
长期以来,演化总被人们认为过程极其缓慢——似乎只有用地质年代来衡量才能看见变化,从恐龙灭绝到哺乳动物崛起、鲸鱼由陆生到水生,这些壮观事件才让我们感受到物种变迁的宏大。但其实,近几十年来的科研工作颠覆了传统观念。越来越多的实验证据显示:只要环境压力足够强烈,许多生物种群会在区区几十年、甚至几年内,表现出显著的新性状,完成“快进版”演化。尤其是在现代社会,人类活动已经成为许多物种演化的加速器。
害虫、杂草与微生物的耐药演化
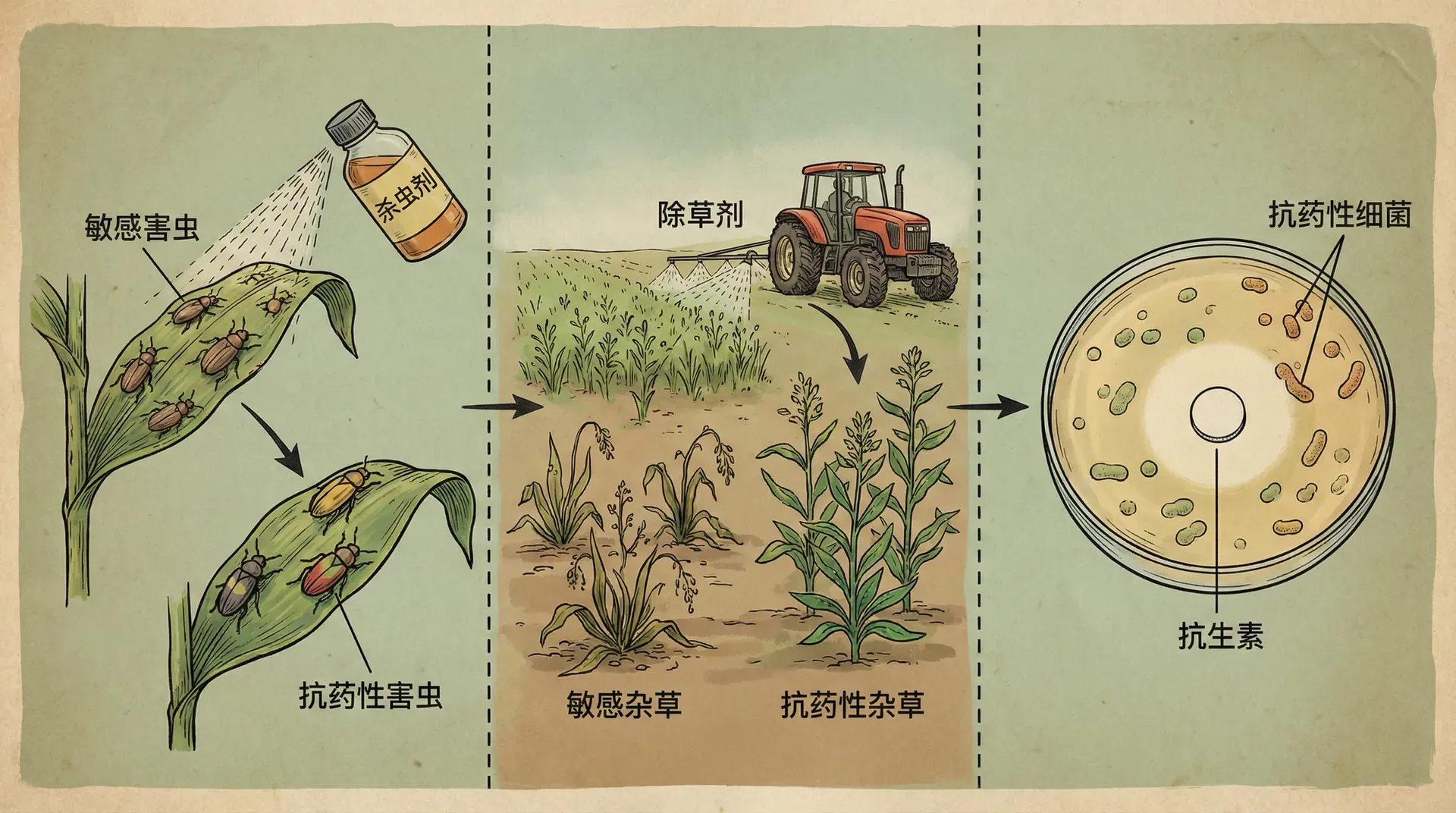
以水稻田里的褐飞虱为例:在中国南方,这种害虫一直困扰着农民。20世纪80年代,科学家研发了新型杀虫剂(如吡虫啉),对褐飞虱的抑制效果极好。然而没过几年,情况急转直下——褐飞虱对吡虫啉的抗药性在多个省份飙升,喷药已经难以奏效。一些试验数据显示,十年间抗药性可以提升数百倍甚至上千倍。
但类似的故事并不只发生在水稻田。我们可以更系统性地梳理一些常见的“快速演化”例子(下表为例):
这些物种通常具备三个共同特征:
- 种群规模极大(基因多样性高)
- 繁殖周期短,一年可繁殖多代
- 受到持续而强烈的人为选择压力(药剂、捕杀等)
譬如说初始时种群只有0.1%个体偶然带有抗药基因,第一次喷药后,99.9%被清除,极少幸存者大量繁殖。下一代抗药比例迅速攀升至10%,继续喷药后该比例又可能升至80%以上,如此几轮筛选后种群“全体”几乎都具备抗药性,传统农药逐渐失效。这个过程可以简化为如下表格:
不同于过去只关注“恐龙灭绝—新物种崛起”这类叙事,现代人眼前每年每季都在发生着“加速版”的演化实例。例如城市蚊子对灭蚊剂耐受力显著增强,非法进口杂草对除草剂的抵抗性突飞猛进,甚至医院细菌对多种消毒剂与抗生素产生多重耐药。
快速演化的关键因素有四:庞大的种群规模、高繁殖率、短世代时间与极强的选择压力。只要这四个条件叠加,自然选择的筛选作用便能在极短时间内将新性状扩展为主流。实际上,科学家已可用实验室“定向进化”方法,在数日或数周内亲自见证新的耐受型细菌或果蝇品系的诞生。
快速演化对种群数量与生态管理的启示
快速演化不仅使生物性状发生突破性变化,更深刻影响了种群数量、波动甚至人类生态治理策略。我们以杂草为例:传统除草剂长期高强度使用,种群迅速演化超级抗性,导致除草剂失效,田间杂草增加,耕作难度反而加大。防治“反被防治误”,经济损失骤增,为此农业领域不得不更新管理思路:比如药剂轮换、化学与生物防治协同、防止单一压力,同时利用“抗性管理”手段延缓演化速度。
生态学的最新研究更指出,快速演化与种群波动、生态系统平衡互为因果:
快演化还广泛影响着自然与人类社会,比如:
- 外来物种(如加拿大一枝黄花)在入侵区迅速适应
- 疾病病原体逃避疫苗的变异
- 城市野生动物对高温、污染和新型食源的适应性变化
现代生态治理不能再简单地将“演化”视为化石堆里的慢镜头。演化本质上是现实世界中时时上演、与种群数量波动反复交错的动态力量。未来可持续的发展与有效控制,必须正视并整合快速演化机制,只有这样才能制定与时俱进、灵活有效的生物管理和生态保护策略。
人类活动引发的演化后果
如果说农药是一种明显的选择压力,那么捕捞和狩猎也是。当我们选择性地移除种群中的某些个体时,就在不知不觉中推动了演化。
长江鱼类的演化响应
长江曾经是世界上最富饶的淡水渔业资源地之一。草鱼、青鱼、鲢鱼、鳙鱼这“四大家鱼”孕育了中国的渔业文明。但从20世纪80年代开始,过度捕捞导致渔业资源急剧下降。令人意外的是,除了数量减少,鱼类本身也在发生变化。
渔业调查发现,同一种鱼的性成熟年龄普遍提前了。比如草鱼,在1970年代,通常需要生长到4-5岁、体重达到3-4公斤才会第一次产卵。但到了2000年代,许多草鱼在2-3岁、体重只有1-2公斤时就开始繁殖了。鱼的平均体长也在缩小,大型个体变得罕见。
这背后的演化逻辑是这样的:捕捞作业往往使用网眼有一定规格的渔网,小鱼能漏网逃生,大鱼却难逃一劫。这意味着,那些能在年轻、体型较小时就繁殖的个体,有更大机会在被捕捞前留下后代。而那些按照“传统节奏”慢慢生长、晚些繁殖的个体,很可能在达到性成熟之前就被捕获了。世代更替之后,早熟、小型化的基因在种群中累积。
图中展示了长江草鱼在几十年间性成熟年龄和平均体长的下降趋势。蓝线表示平均性成熟年龄从4.5岁降到2.5岁左右,红线表示平均体长从55厘米降到36厘米。这种变化速度之快,说明捕捞压力作为选择力量是何等强大。
演化稳定的捕捞策略
这些演化变化对渔业本身有什么影响?当鱼类提前成熟,它们会把更多能量投入繁殖而不是生长。结果是,虽然鱼的数量可能不会立刻大幅下降,但每条鱼都变小了,总产量实际上在减少。渔民需要捕捉更多的鱼才能获得同样的收益,这进一步加剧了捕捞压力,形成恶性循环。
那么,有没有一种捕捞方式,既能保证长期产量,又不会驱动不利的演化?有研究提出了“演化稳定最优捕捞策略”的概念。简单说,就是要让捕捞方式和鱼类的生活史相匹配,避免过度选择某个年龄段或体型的个体。
比如,传统上在产卵场捕鱼可能比在觅食地捕鱼更好。因为在产卵场,捕捞压力对所有成鱼都相对均等,不会特别偏向某个年龄。而在觅食地,如果持续移除大鱼,就会选择出早熟的小型鱼。
中国从2021年开始在长江流域实施十年禁渔,这是一个极其重要的举措。这不仅给鱼类种群数量恢复提供了时间,也给演化一个“喘息”的机会。在没有捕捞压力的情况下,那些生长缓慢但最终体型较大、繁殖力更强的基因型可能会重新获得优势。当然,这个过程需要时间,可能需要几代鱼才能看到明显的逆转。
适应度地形的复杂性
现在让我们深入思考一个更理论性的问题:演化总是朝着“更好”的方向进行吗?
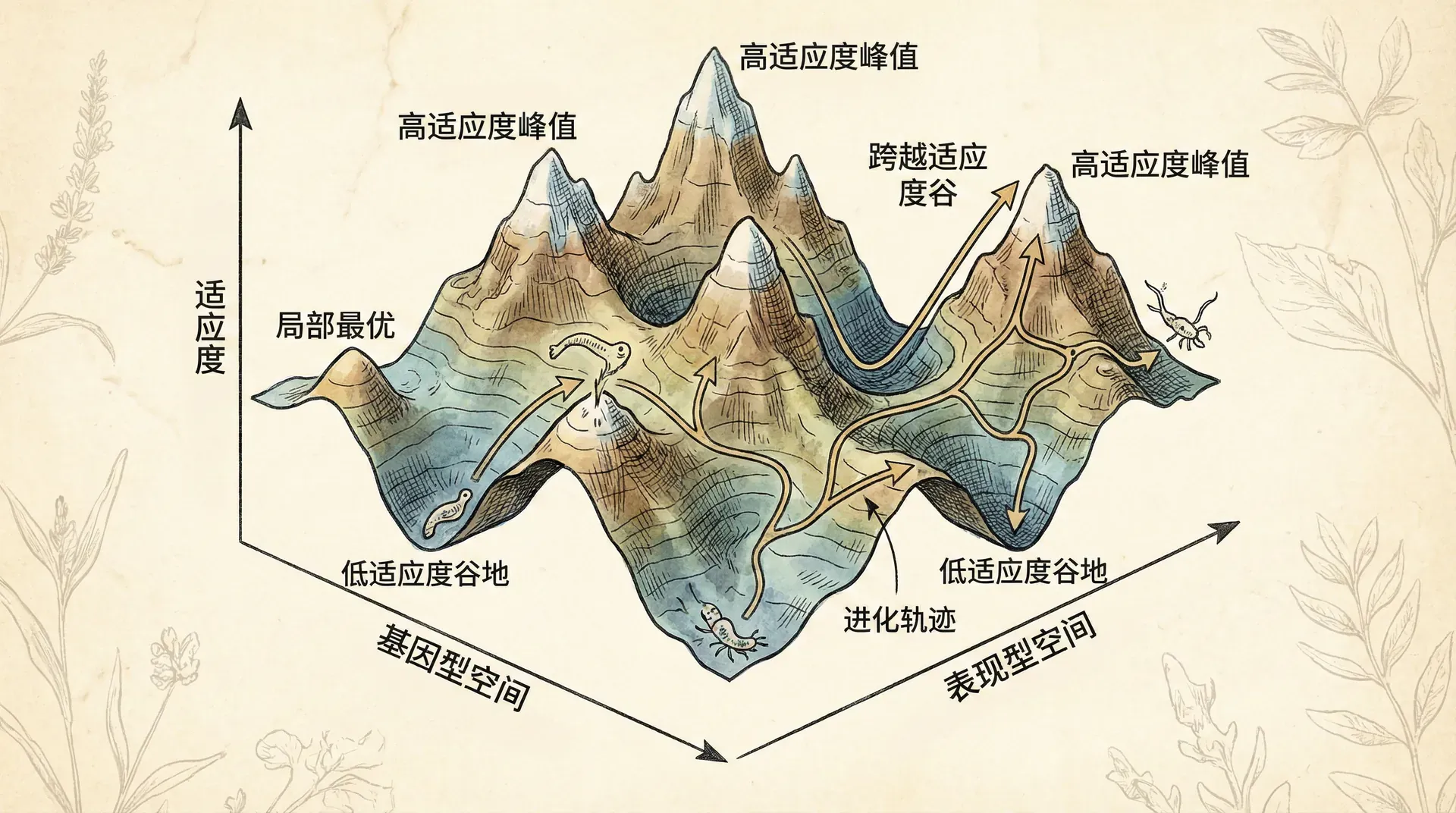
演化不总是爬坡
传统的演化观点常常用“适应度景观”来比喻:把生物的不同特征想象成一个起伏的地形,高处代表适应度高,低处代表适应度低。演化就像爬山,种群通过自然选择不断向高处移动,最终到达山峰。
但现实比这复杂得多。这个比喻的问题在于,它假设地形是固定的。实际上,当种群演化时,个体之间的相互作用也在改变,适应度景观本身就在变化。
例如,在一个湖泊里,鱼类的体型影响它们的竞争能力和食物选择。假设最初大型个体有优势,演化推动种群向大型化发展。但当大多数鱼都变大之后,它们之间的竞争变得激烈,对大型猎物的需求超过了供给。这时候,中小型个体反而可能因为能利用不同的食物资源而获得优势。
这种情况下,演化可能会到达一个奇怪的点:这个点本身的适应度低于周围的很多策略,但演化过程恰好把种群带到了这里。一旦到达,周围所有方向的突变个体都比现在的主流类型更有优势。这就像爬到了山顶,突然发现山顶塌陷成了盆地。
演化分支与多样性
当到达这样的“适应度谷底”时,会发生什么?理论上,任何方向的变异都会受到选择的青睐。这意味着选择是发散的——不同方向的变异可以同时在种群中存在。如果两种不同的变异都能稳定共存,就形成了多态性。极端情况下,这可能导致一个种群分化成两个不同的类型,甚至最终形成不同的物种。
这个理论被称为“适应性动力学”,它考虑了种群密度、个体间的竞争等生态因素如何影响演化方向。这个视角很重要,因为它解释了为什么我们在自然界中看到那么多的多态现象——同一个种群内不同的体型策略、不同的繁殖时间、不同的扩散倾向等等。
在一个森林里,树木竞争阳光。一开始可能高大的树有优势。但当森林中都是高大的树时,林下形成了一个不同的生态位。这时候,能够在阴暗环境中生存、利用不同资源的矮小灌木也有了生存空间。演化不再是单纯的“越高越好”,而是分化出了“高大乔木”和"耐阴灌木"两种策略。
将种群动态纳入演化思考,能帮助我们理解生物多样性的起源。演化不是简单的优胜劣汰,而是一个会产生分支、创造新可能性的动态过程。
总结
回到最初的问题:演化如何影响种群数量,种群动态又如何反过来影响演化?两者密不可分。比如捕食者数量增多,可能促使猎物演化出更强防御或更快繁殖速度,反过来,猎物变化也会推动捕食者作出适应调整,形成复杂的反馈回路。个体的适应行为(如迁徙、改变繁殖时间等)可以快速响应环境变化,而演化也能够在可观测的时间范围内影响群体结构、数量和生态功能。更进一步,生态互动(如竞争、捕食、共生)常推动新物种的产生和多样性的增加,使演化过程充满变数。
这些原理已深刻影响实际应用:医学中重视“达尔文医学”,避免抗药性快速产生;渔业管理倡导将生态与演化视角结合,防止捕捞诱导有害性状;农业防治强调抗性管理,遏制害虫耐药性进化。
融合演化与生态学,为我们更科学地理解和管理自然系统提供了基础。生命不是被动应对环境,而是在遗传变异和选择下主动重塑生态,而环境本身也在生物活动影响下不断演变。只有双重视角,才能应对健康、保护与可持续发展的挑战,做出明智决策。