生物扩散与休眠的进化
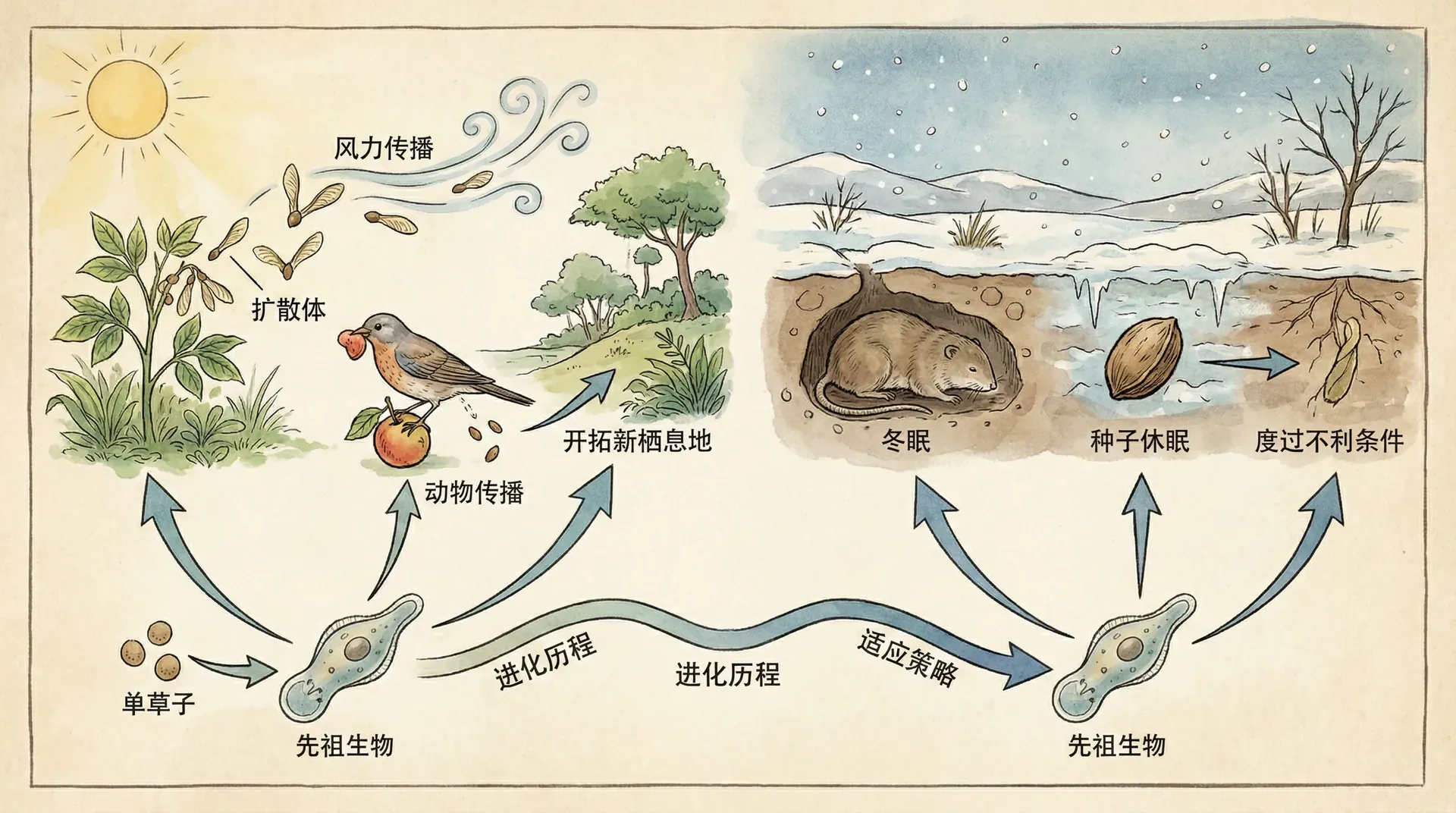
自然界中,生物应对环境不确定性的策略丰富多样。秋季时候鸟远渡重洋,面对风暴、饥饿与敌害,迁徙之路充满风险,却也为种群争取到了更多生存可能。而在干旱沙漠里,沙米种子常常长时间休眠,静待理想时机再破土发芽。无论是主动迁徙还是被动等待,这些策略背后都体现了生物为分散风险、适应环境变化所做的进化选择。
表面上,迁徙和休眠是两种完全不同的生命现象,一个选择“走出去”,一个选择“留得住”。实质上,它们的共同点在于都避免了“把所有希望押在一个时空”,从而增加了物种持续繁衍的机会。不同生物甚至在同一物种内,也常结合使用这些策略,比如鱼类产下可耐干旱的卵,真菌孢子随风扩散。
总的来看,扩散和休眠不仅仅是对突发灾害的被动应对,更是生物在进化中精心打磨、应对不确定性的核心智慧——只有通过“分散风险”,生命才有更大概率在不断变化的世界中延续下去。
扩散行为的进化难题
从群体选择到个体选择
20世纪60年代,英国生物学家温-爱德华兹提出了一个大胆的假说:动物通过扩散调节种群数量,避免过度繁殖导致资源耗尽。那些“不自私”的个体主动离开,为留下的同伴留出生存空间,最终这样的种群得以延续。这个理论听起来很美好,但存在一个致命缺陷。
如果整个种群都遵循“为集体牺牲”的策略,那么一个“投机者”突然出现了——它不参与扩散,只享受别人扩散带来的好处。这个投机者的生存率更高,后代也更多。经过几代繁衍,投机者的基因会迅速扩散,最终取代那些“无私”的个体。这就是为什么单纯的群体选择理论站不住脚。
进化并不总是选择对种群最有利的策略,而是选择对个体最有利的策略。这意味着有时候个体的最优选择,反而可能增加种群灭绝的风险。
进化稳定策略的诞生
那么,扩散行为到底是如何进化出来的?答案在于“进化稳定策略”这个概念。简单说,这是一种一旦被种群采纳,就无法被其他策略入侵的行为模式。
1977年,汉密尔顿和梅提出了一个简洁而强大的数学模型。他们设想一个简化的世界:环境由许多相同的栖息地斑块组成,每个斑块住着一个成年个体。每年,成年个体死亡,它们的后代竞争这些空出的斑块。一部分后代选择扩散到其他斑块,另一部分留在原地。
模型得出的结论令人惊讶:最优扩散率为 ,其中 是扩散者的存活率。这意味着即使扩散的死亡率极高(比如存活率只有10%),仍然应该有超过50%的个体选择扩散!
为什么会这样?关键在于亲缘竞争。留在原地的后代会和自己的兄弟姐妹竞争,它们携带相同的基因。而扩散的后代虽然面临死亡风险,但一旦成功,就能占领新的斑块,与非亲缘个体竞争。从基因传递的角度看,让一部分后代扩散,实际上是一种明智的"投资组合"策略。
以下是扩散率与存活率的关系:
这个模型还揭示了一个重要现象:从个体角度的最优扩散率,往往高于从种群角度的最优值。换句话说,自然选择会推动扩散率超过最有利于种群生存的水平。这再次说明,进化优化的是个体适应度,而非群体利益。
环境变化
时间的不确定性
如果环境是恒定的,今年适合生存的地方明年依然适合,那么扩散的意义就大打折扣。但现实世界充满变化。一个今年风调雨顺的草原,明年可能因干旱变得不适宜生存;一棵今年结实累累的树,明年可能遭遇病虫害。
时间上的变化促进了扩散行为的进化。这就像投资理财中的“不要把鸡蛋放在一个篮子里”。把后代分散到不同的地方,就是在对冲环境波动的风险。即使父母所在的栖息地今年条件很好,也不能保证明年依然如此。
不同类型的时间变异及其对扩散行为的影响:
内蒙古草原上的东亚飞蝗就是典型例子。在降雨充足的年份,草原植被茂盛,蝗虫密度较低,大多数个体翅膀较短,不太飞行。但一旦遭遇干旱,植被稀疏,蝗虫密度激增,新一代蝗虫就会发育出长翅膀,形成蝗群迁飞。这种可塑性正是对环境时间变异的适应。
云南的部分竹种会在几十年后集体开花死亡,这让依赖竹林的动物(如大熊猫)必须迁移到其他竹林。这种现象被称为“扬森-康奈尔假说”:父母周围积累的天敌和病原体,使得原地对后代不再适宜,从而强烈选择扩散行为。
空间的异质性
与时间变异相反,空间上的差异往往抑制扩散。如果栖息地之间质量差异很大,那么待在好地方的个体如果扩散,更可能落到质量较差的地方,得不偿失。
中国海岛上的生物为这个理论提供了有力证据。舟山群岛、南海诸岛上的许多鸟类和昆虫,相比大陆同类,飞行能力明显退化。比如,台湾岛上有多种特有的无翅昆虫,它们的大陆近缘种都具有完整的翅膀。原因很简单:在岛屿上,飞行能力强的个体更容易被风吹到海里,留下来的都是那些飞不动或者不爱飞的。经过无数代选择,岛屿物种的扩散能力就这样退化了。
青藏高原的特有物种也呈现类似模式。高原环境相对稳定,但被周围的低海拔地区“包围”。对高原物种来说,向外扩散意味着进入低海拔的不适宜环境。长期下来,许多高原特有种的扩散能力都比较弱。
下方对比了不同栖息地类型中物种的扩散倾向:
从图中可以清晰看到,临时性强、变化快的栖息地(如农田)中的物种更倾向于发展强扩散能力,而稳定、孤立的栖息地(如海岛、洞穴)中的物种扩散能力普遍较弱。
性别与扩散
在许多哺乳动物中,雄性比雌性更爱“离家出走”。这是为什么?答案与交配制度有关。
在一夫多妻制的物种中,雄性之间为了交配权展开激烈竞争,而雌性主要竞争食物等资源。如果雄性留在出生地,就会与自己的兄弟争夺交配机会,这种竞争尤其激烈,因为它们携带相似的基因。而扩散到新地方,虽然仍要竞争,但对手是非亲缘个体,从基因传递角度更有利。同时,雄性扩散降低了近亲繁殖的风险,让留守的雌性受益。
但在一夫一妻制且雄性需要占据领地资源才能吸引配偶的物种中,情况可能逆转。此时雌性之间对资源的竞争可能更激烈,反而是雌性更倾向于扩散。
扩散行为的性别差异不是随机的,而是由亲缘竞争的强度和交配制度共同决定的。这个原理在从昆虫到哺乳动物的广泛类群中都得到了验证。
来自野外的证据
昆虫的飞行能力
理论预测是一回事,自然界的真实情况又是另一回事。那么,野外观察支持这些理论吗?
对中国昆虫的研究提供了丰富的证据。在华北平原的农田生态系统中,研究者调查了数十种叶蝉和飞虱。这些昆虫的栖息地——农作物——每年都要收割重种,是典型的“临时性栖息地”。调查发现,超过80%的农田叶蝉物种具有完全发育的翅膀和强飞行能力。相比之下,森林中的同类昆虫,无翅或短翅类型的比例明显更高。
更有趣的是,一些昆虫物种内部出现了“翅二型”现象:同一个物种的不同个体,有的长翅有的短翅。这种可塑性让它们能根据环境条件灵活调整策略。比如,褐飞虱在食物充足、密度较低时倾向于发育短翅,节省能量用于繁殖;而在食物不足、密度过高时,新一代主要发育长翅,准备迁飞到新的稻田。
蜻蜓的迁飞模式也验证了理论预测。中国有记录的迁飞性蜻蜓,如黄蜻和蓝额疏脉蜻,主要栖息在临时性水体,如季节性湖泊和水坑。而那些生活在溪流、永久性池塘中的蜻蜓物种,很少进行长距离迁飞。
植物的种子传播
植物无法移动,但它们的种子可以旅行。中国植物区系为研究扩散进化提供了理想的自然实验室。
蒲公英属植物在中国分布广泛,从东部平原到青藏高原都有分布。研究发现,低海拔地区的蒲公英种子冠毛(就是那些白色的“小伞”)发达,风一吹就能飞出上百米。但在青藏高原上的高山蒲公英,冠毛明显退化,有些种类甚至几乎没有冠毛。这符合空间异质性理论:高原环境孤立而特殊,种子飞出高原很难存活,自然选择反而青睐那些“待在原地”的个体。
海南岛上的植物也展现出类似模式。与大陆近缘种相比,许多海南特有植物的种子失去了翅、毛等助飞结构,变得更重、更难被风带走。这是典型的岛屿化效应:强扩散能力的种子容易被吹到海里,留下的都是扩散能力弱的后代。
下图是栖息地稳定性与扩散能力的关系:
数据清楚地显示:栖息地越临时、变化越快,物种就越倾向于发展强大的扩散能力。这条规律在昆虫和植物中都成立。
分布区边缘的快速进化
近年来气候变暖,许多物种的分布区正在向北、向高海拔扩张。这为研究扩散进化提供了难得的“实时”机会。
黑斑蛱蝶原本分布在中国中部和南部,但最近30年逐渐向北扩散,现在已经在河北北部建立了种群。对不同地区种群的对比研究发现,分布区北缘的蝴蝶胸部肌肉更发达(那是飞行的动力器官),飞行能力更强,但产卵量较少。这说明在扩张前沿,自然选择正在快速塑造更强的扩散能力,代价是繁殖力下降。
这个发现让人担忧:如果扩张中的物种会进化出更强扩散能力,那些因栖息地破碎化而萎缩的物种会怎样?理论预测它们可能进化出更弱的扩散能力,就像岛屿物种一样。这意味着它们会更难重新扩散到其他适宜栖息地,进一步增加灭绝风险。这种“进化自杀”现象,是保护生物学必须认真对待的问题。
种子的三种策略

休眠的代价与收益
说完扩散,我们再来看另一个有趣的现象:种子休眠。许多植物的种子成熟后并不立即萌发,而是在土壤中“睡上一觉”,甚至一睡就是好几年。
休眠显然有代价。首先,种子在土壤中每年都有一定的死亡率,可能被真菌感染、被动物吃掉、或者失去活力。其次,在种群数量增长的阶段,早繁殖意味着多繁殖——你的后代能经历更多代,基因传播得更快。休眠延迟了繁殖,从这个角度看是不利的。
但休眠也有明显的好处:它是时间维度上的“风险分散”。今年可能是干旱年,全部萌发的种子都会死亡;但如果一部分种子休眠,等到明年雨水充足时再萌发,就能逃过这一劫。这和扩散在空间上分散风险的逻辑完全一样,只不过休眠是在时间上下注。
中国西北干旱区的植物普遍具有较强的种子休眠性。对索诺兰沙漠植物的研究(虽然不在中国,但原理相通)表明,降雨量年际波动越大的地区,植物的种子休眠比例越高。这符合理论预测:环境越不可预测,“押注”未来的策略就越有价值。
另一个促进休眠的因素是亲缘竞争。如果种子全部在父母植株附近萌发,兄弟姐妹之间的竞争会非常激烈。通过休眠,一部分种子错开竞争时间,可以提高家族整体的成功率。
种子大小的意义
除了扩散和休眠,种子还有第三个选项:改变大小。大种子储存更多养分,幼苗能更好地应对不利环境,比如干旱、贫瘠土壤、或浓密的竞争者。但大种子意味着母株必须投入更多资源,同样的资源只能生产更少的种子。
那么,一个植物应该生产少量大种子,还是大量小种子?答案取决于环境条件。在竞争激烈、资源贫瘠的环境中,大种子有优势。但在开阔、资源丰富的环境中,小种子数量多反而更有利。
三角权衡关系
现在问题变得复杂了:植物面临三个选择——扩散、休眠、还是增大种子?有趣的是,这三个策略实际上在功能上相互替代。
扩散是在空间上逃避不利环境;休眠是在时间上逃避不利环境;而增大种子是在原地“硬扛”不利环境。如果把资源投入到扩散(比如发育更大的翅或冠毛),就没有足够资源用于休眠机制或增大种子。这就是所谓的“生活史权衡”。
1988年,维纳布尔和布朗发表了一篇经典论文,从理论上分析了这三个性状如何共同进化。他们的模型预测:
为便于比较不同生境类型植物在扩散和休眠策略上的差异,我们可以用以下信息概括:
可以看出:
- 沙漠植物偏向于时间上的赌注——高比例休眠以抵御年际极端干旱;
- 湿地植物则倾向于空间上的赌注——扩散距离远,抓住短暂适宜条件;
- 森林植物相对均衡,休眠和扩散适中,但更注重提升种子本身的竞争力。
这种权衡反映出不同生态环境下,植物通过分配有限资源,在“扩散”、“休眠”和“种子大小”三者之间进行最优配置,以应对各自独特的生存挑战。
真实世界的验证
英国植物区系的数据为这个理论提供了有力支持。研究者测量了数百种植物的种子质量、扩散距离和休眠倾向,发现这三个性状确实存在显著的负相关:扩散能力强的物种,种子往往较小且休眠性较弱;种子大的物种,扩散距离短且休眠比例低;休眠性强的物种,扩散距离和种子大小都偏小。
更直接的证据来自岛屿植物。许多岛屿特有植物不仅扩散能力退化,种子还明显变大。塞舌尔群岛的海椰子就是极端案例——它有世界上最大的种子(重达20多公斤),几乎完全不扩散,也没有明显的休眠期。这完美诠释了权衡关系:当扩散没有意义时(周围都是海洋),植物把全部资源投入到增大种子上,让幼苗拥有巨大的竞争优势。
中国的实例同样丰富。比较长江流域和海南岛的同属植物,岛屿类型的种子平均重量增加约40%,而风媒传播结构的发生率降低约35%。这再次验证了理论预测。
下面总结了不同环境因素如何影响这三个性状:
种子的扩散、休眠和大小并非独立演化,而是在资源限制下的联合优化。环境的细微变化会同时重塑这三个性状,形成不同的生活史策略组合。
总结
通过分析扩散和休眠的进化,我们看到了自然选择的复杂性和精妙性。这些行为模式不是随意出现的,而是在特定环境压力下,经过无数代优化的结果。
从汉密尔顿和梅的简洁模型,到维纳布尔和布朗的多性状权衡理论,进化生物学家们用数学语言刻画出生命策略的内在逻辑。而大自然用千百万个物种的真实表现,验证了这些理论的预测。从内蒙古草原的飞蝗,到青藏高原的特有植物,从候鸟的迁徙路线,到沙漠种子的休眠周期,都能找到进化理论的印记。
这些理解不仅满足了我们的求知欲,还有重要的实践意义。当我们进行生态保护时,必须意识到物种会在我们的干预下继续进化。栖息地破碎化可能导致物种扩散能力退化,进入“进化陷阱”;气候变化会重塑扩散和休眠的最优策略;人工栖息地的创建需要考虑物种的扩散能力和生活史策略。
理解生命的进化逻辑,不仅能帮助我们预测物种会如何响应环境变化,更能指导我们设计更有效的保护策略。生态保护不是简单地“冻结”当前状态,而是要为进化过程留出空间,让生命的智慧继续延续。
扩散与休眠,这两个看似简单的生命策略,背后蕴含着如此丰富的进化智慧。它们提醒我们,生命的每一个决策都是在复杂的约束和权衡中做出的最优选择。在不断变化的环境中,“旅行者、留守者与沉睡者”各有各的生存之道,共同编织出生命世界的多样性和韧性。