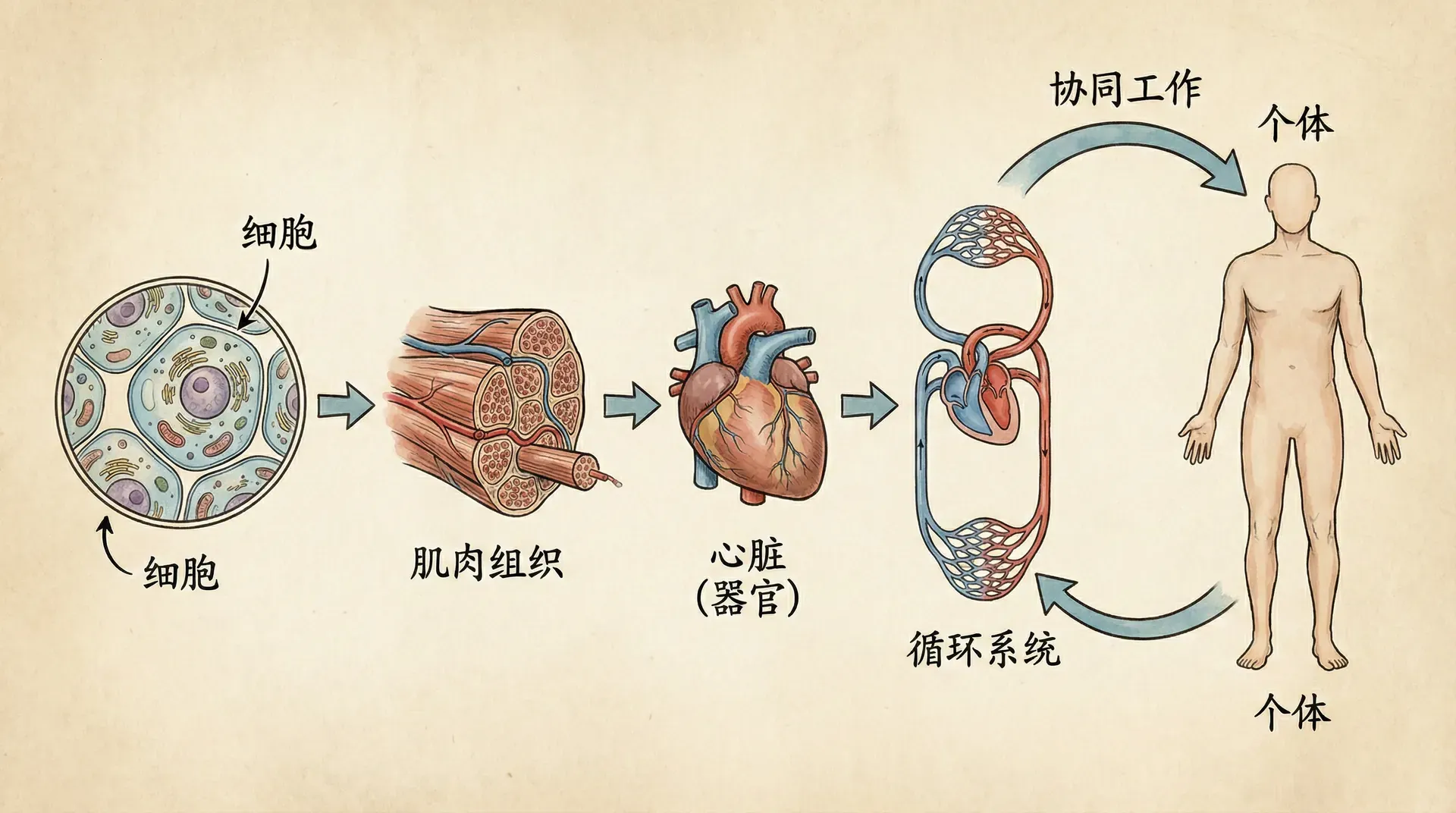
生物组织的层次与协作
当我们走进一片竹林,眼前是一片郁郁葱葱、层层叠叠的竹竿,这些竹竿仿佛在同一个节奏下一起生长,蔓延成绿色的海洋。这时你可能会产生疑问:这片竹林,到底算是一个生物,还是无数个彼此独立的生物个体?表面上看,每根竹竿都很独立,它们各自生长、发育、高高挺立;但如果你俯身观察或者试图挖开地表,你会发现:所有这些竹竿其实都通过地下的竹鞭(地下茎)彼此联结,共享着同一套根系和水分养分。表面上彼此分离、实则暗中相连,这种现象令人不禁思考:究竟该如何界定“一个生物个体”?这个看似朴素的问题,实际上触及了生物学中最基础但又异常复杂的议题——如何定义“个体”与“整体”。
我们平常理解里,往往把生物看作清楚分明的“个体”:每个人是一个个体,一只狗或者一棵大树,我们本能地认为它们就是完整、独立的生命单位。然而,当代生物学的发展早已打破了这种黑白分明的印象。在自然界中,既有单独生活的孤生细胞,也有成千上万细胞组成的大型多细胞生物,甚至还有复杂到令人惊叹的集体生命现象,比如真菌菌丝网络、蚂蚁社会、珊瑚礁群落等。
因此,定义“生物个体”远不像想象中那么简单。这不仅涉及到我们观测生物的角度,也关乎生物自身是如何维系生存和进行繁殖的。接下来,我们将通过更多例子逐步揭开生物组织层次与协作的多样性,让大家认识到“个体”的边界其实充满弹性与活力。
重新认识生物个体
我们平时觉得区分一个生物个体很容易,但仔细观察就会发现许多令人困惑的现象。
在长江流域的池塘里,水螅通过出芽方式繁殖。母体身上长出小芽,这个芽逐渐发育出触手和消化腔,最终可能脱离母体独立生活,也可能一直连着。那么在芽还没完全分离时,它算是母体的一部分,还是已经成为一个新个体了?
一些千年银杏能够通过根系萌发新芽,在原树周围长出一圈“子树”。这些子树与母树基因完全相同,根系相连,营养共享。从遗传角度看,它们是同一个个体的克隆;但从功能角度看,每棵树又能独立进行光合作用和生长。
生物个体的界限并非总是清晰可辨。在自然界中,生物体的边界往往取决于我们从什么角度去观察——是从遗传物质、物理结构,还是从功能协作的角度。
为了更好地理解这个问题,生物学家提出了“自我维持系统”这个概念。例如,一支蜡烛的火焰。火焰看起来稳定存在,但实际上构成火焰的物质每时每刻都在变化——蜡不断融化、燃烧,空气不断流入,产物不断排出。火焰之所以能维持,是因为它形成了一个动态平衡:燃烧产生的热量维持了温度,高温又促进了持续的燃烧反应。
生物体也是类似的自我维持系统,但比火焰复杂得多。一个细胞需要不断从外界获取营养物质,进行代谢反应,产生能量,合成自身组分,排出废物,同时还要调控这整个过程。这种能够维持自身存在、抵抗外界干扰的能力,是生物区别于非生物的关键特征。
对比不同生物个体在边界定义上的差异:
判断生物个体的标准可以多样化,但核心在于考察该系统是否能够自我维持、自我调节,以及在多大程度上依赖外部条件。
下方展示了不同生物组织形式的复杂度差异:
从图中可以看出,随着组织复杂度提升,生物系统的自我维持能力和功能分化程度都在显著增强。原核细胞(如细菌)虽然结构简单,但已经具备了相当强的自我维持能力。而到了复杂多细胞生物阶段,不同细胞类型高度分化,协作完成更复杂的生命活动。
单细胞生物的精妙调控
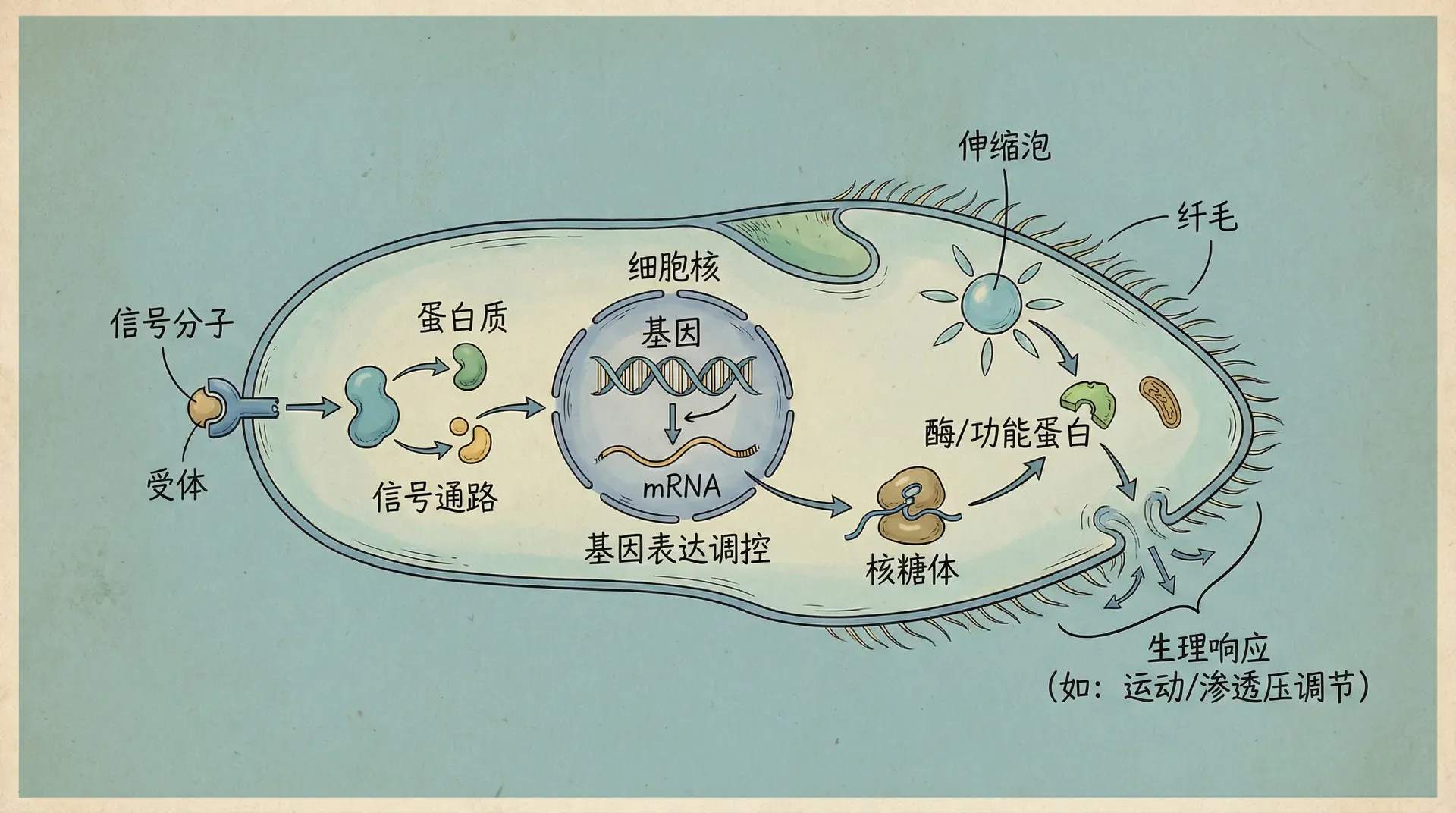
单细胞生物虽然体积微小,但展现出的“生存智慧”却令人惊叹。它们远非一颗简单的“活着的颗粒”,而是高度复杂、动态自组织的生命系统。单细胞生物凭借巧妙的分子机制,不仅能感知环境、处理信息,还能做出灵活的行为决策,这些机制是生命起源和演化不可或缺的基础。
让我们以变形虫(Amoeba) 和大肠杆菌(E. coli) 为例:
大肠杆菌具有极为灵敏的化学感受能力。比如在污水处理厂中,大肠杆菌能够在营养稀缺的水体内执行“翻滚运动”:通过快速、随机地改变游动方向,增大探索空间。如果细胞膜上的趋化受体检测到葡萄糖等营养物质浓度提升,细胞可以极速(通常不到一秒)切换到“定向游动”,朝目标前进,展现出灵活应变的本领。
趋化性运动依赖于一个环环相扣的信号网络,包括受体、信号传导系统和运动装置。
信号传导过程中的“双组分系统”尤为关键。受体捕捉信号后,将信号以磷酸基团的形式递进传递,层层放大,最终让鞭毛马达快速、精准地响应。这种级联机制不仅让细胞能实时判断周围环境,还实现了多重信号整合和优先级判断。
更有趣的是,大肠杆菌等单细胞生物还拥有 “分子记忆” 能力。举例来说,细胞可通过蛋白质的甲基化/去甲基化变化,记住过去的化学环境,从而将当前检测到的信号与过去做差异比较,借此决定是否“继续前进”或“改变方向”。这类分子记忆虽简单,却极大提升了单细胞应对波动环境的适应性与“决策力”。
这些机制共同构成了一个完整的 “感知—处理—响应”闭环。即便没有大脑或者神经网络,单细胞生物依然能展现令人惊叹的复杂行为。这种分子层面的“智能”,是单细胞生物能够成功生存和持续进化的根本。
细菌的群体协作
当无数细菌汇集时,便会催生全新层次的集体行为,这是从“独自生存”进化到“合作生存”的重要跳板。
例如在自然界或人工环境中,铜绿假单胞菌(Pseudomonas aeruginosa)、乳酸菌等细菌常常分泌黏液,在固体表面形成生物膜(biofilm)。生物膜不仅是细菌消极聚集形成的产物,更是通过复杂的信号交流实现行为协调的“微型社会”。下表展示了常见细菌生物膜的特征:
以“生物膜”为例,生物膜广泛存在于污水处理系统(如活性污泥法)、人体口腔的牙菌斑、医疗器械表面等环境。生物膜里的细菌不仅能共享资源、抵抗威胁,还会根据整体情况调整单体行为。例如,营养匮乏时,细菌可同步滑动作为集体向新区域迁移。
生物膜在污水处理、环境修复和医疗防护等领域有重要作用。例如,活性污泥法能够依靠生物膜中的细菌共同分解有机污染物,大幅提升处理效率。
这些“城市化”的协作由所谓的群体感应(quorum sensing)机制驱动。细菌会分泌特殊的信号分子(如自诱导因子autoinducer),并能侦测这些分子的浓度,进而估算周围同类个体的数量。一旦感应到菌群密度超越某个阈值,细菌便会同时激活一系列基因,实现统一行为调整。例如,剧毒绿脓杆菌在密集状态下才会集体爆发致病力。
此外,一些细菌(例如社会性黏菌、放线菌等)在极端环境下,会组装成“子实体”(fruiting body)结构。部分细胞分化成孢子以便延续后代,其他细胞则牺牲成为支撑结构。这种协作虽已涉及功能分化与团队作业,但一般是短暂、可逆的应急机制。环境一旦改善,孢子萌发,各自又回归单细胞状态。因此,遗传和代谢独立性仍是在此协作中的主导。
本质区别总结表:
简而言之,细菌群体合作为生命组织进化提供了无限可能。尽管这些协作远不及多细胞生物的整合和分化程度,但已在组织、通信和功能层面奠定了进化的台阶,是迈向高级生命不可或缺的阶段。
真正的多细胞组织
从生物演化的角度来看,生命体由单个细胞逐步进化到复杂的多细胞系统,期间经历了信息交流、功能分化、结构整合等多个关键阶段。多细胞组织的出现,是生物体在复杂度和功能性上实现跃迁的重要标志。真正意义上的多细胞生物,不仅包含多种功能不同的细胞类型,而且这些细胞通过发达的联系和信号调控形成稳定、协作、不可分割的有机整体,从而支撑起高度复杂的生命活动。理解多细胞组织的演化及其整合度变化,有助于揭示生命多样性与高级功能的根本来源。
生物体的组织整合度经历了由低到高的递进过程,呈现出五个清晰阶段:
-
单一细胞阶段(细菌):如大肠杆菌,只包含单一类型的细胞,虽然可以以群体形式存在(如生物膜),但每个细胞在结构和功能上彼此独立,整合度最低。
-
松散群体阶段(生物膜细胞):如污水系统的活性污泥细菌,虽然个体聚集形成生物膜,通过分泌物物理连接,但细胞类型依旧单一,群体协调有限,只能通过“群体感应”实现简单通信,整合度较低。
-
暂时分化阶段(黏菌子实体):如黏菌,在特定条件下出现两种功能分化的细胞——孢子与支撑细胞,实现协作分工,但这种分化和结构仅在环境压力下短暂存在,分化状态可逆,整合度中等。
-
简单多细胞阶段(团藻体细胞/生殖细胞):如团藻,由体细胞和生殖细胞两种类型组成,分工永久固定,结构上由胞外基质和细胞质桥连接,各细胞任务明确,整合度显著上升,但生殖细胞仍可脱离整体独立生活,个体整合有限。
-
高度分化阶段(多细胞动物):如有神经、肌肉、消化等专职细胞的多细胞动物,细胞分工高度专业化,各功能不可逆转,失去个体部分便无法存活,整体行为远超各部分简单叠加,系统整合度达到顶峰。
以团藻为例,其体细胞和生殖细胞的功能分工是发育早期便决定的。体细胞负责运动与感光,生殖细胞专司繁殖,两者功能不可互换。但根据图表的数据和分层,团藻的结构整合度虽然已达到新台阶,仍明显低于神经动物等更高级多细胞生物。此外,团藻生殖细胞在分离后依然有能力单独生长,体现了整合度的有限性。
下方图表展示了各阶段细胞类型及其对应的组织整合度,滑动选择不同演化阶段,可直观查看对应的细胞类型描述和整合度变化:
综上可见,组织整合度的提升呈阶梯状,在关键的结构和分化节点出现明显跃升。图中60%至95%的巨大跃迁,反映了从简单多细胞(如团藻)向复杂多细胞动物(如拥有神经和肌肉等专职细胞的后生动物)的质变。这一跃升标志着生物个体从“松散联盟”进化为真正不可分割、协同一体的高度整合生命系统。
神经系统
在复杂多细胞生物的演化历程中,神经系统的出现无疑是最具革命性的突破之一。它极大提升了个体内部各细胞之间的整合与协作能力,让生命体能够高效应对不断变化的环境。动物世界的进化,正是沿着神经系统越来越复杂和高效的方向不断前进。
以海洋中的水母和珊瑚虫(如水螅)为例,这些生物看似柔软而原始,却完成了从松散细胞集合体向高度整合个体的飞跃。与团藻那样的简单多细胞生物不同,水母体内分布着由神经元构成的网络系统,如同一张信息“渔网”,贯穿全身,成为信号传递的主干道。这种分布式神经系统没有一个明确的“中心大脑”,但已经可以感知光信号、机械刺激,并能通过网络将不同区域的信息加以整合,实现对全身的快速协调和反应。
我们可以将神经系统的分化整合度通过下表对比:
在水母的游动、捕食与逃逸等行为中,神经网络起到了至关重要的作用。例如,感光细胞检测到环境变化,通过神经元快速将信号传递至相应肌肉,实现身体边缘同步收缩,产生优雅或有力的运动。正是这种分布式的信息整合与高效响应,让水母能根据情境灵活切换生存策略,比如捕食平稳时舒缓运动,遇敌时急促逃逸。
神经系统的本质,是把成百上千的细胞与组织整合为一个统一的信息处理和决策响应整体,让真正意义上的“生物个体”得以诞生。此时,局部的功能不能再脱离整体单独存在,各部分协作成为不可分割的一体。
随着进化深入,神经系统开始出现集成中心。比如蚯蚓等环节动物拥有原始的脑节,集中处理信息,向更高效的“团队协作”迈进。脊椎动物则发展出结构复杂的脑,由各种中枢统领运动、行为、情绪和生理状态。例如,哺乳动物的下丘脑-垂体轴能对体温、代谢、水盐平衡等多项功能进行综合调节。
这样,动物的“生存”(如内部代谢)和“行为”(环境互动)通过神经网络被有机结合。哪怕是结构最简单的水螅或水母,一旦失去关键部分也将丧失协同功能,危及整体生命。
因此,从海蜇、蜜蜂到大熊猫,这些生物之所以可称为“个体”,正是因为它们的局部结构已高度分化且彼此依存,脱离整体就难以独立生存。它们的统一表型和行为,是协同而非简单的局部叠加。
共生

除了单一物种内部的高效整合,生命世界还发展出跨物种的复杂协作——也就是“共生”现象。共生不仅提升了生命系统的整合度,更大幅拓展了生态与功能的边界。
在我国南方水稻田常见的苔草(豆科)与根瘤菌共生是一种典型现象。根瘤菌“入侵”植物根须,在根部形成专门结构,帮助植物固氮,从空气中获取氮源;植物则以光合作用产物“回馈”根瘤菌。这种协作让双方在贫瘠土地上都能存活,彼此离不开。
从进化角度看,线粒体和叶绿体——这些真核细胞里必不可少的细胞器,本身就是“微生物入侵”后长期共生的产物。它们早已与宿主细胞高度融合,形成立体功能整体。
另一个案例是白蚁和肠道微生物的“超有机体”。在南方的白蚁巢穴,白蚁体内居住着丰富细菌、原生动物等微生物,合力分解纤维素使白蚁能汲取养分。如果用抗生素清理微生物,即使白蚁还会觅食,最终也会因无法消化而死亡。许多牛羊等反刍动物体内,也依赖微生物消化草料。
常见共生和整合关系的比较:
我们会发现随着协作层次加深,从松散聚集到神经整合再到共生一体化,不同“协作单元”的独立性逐步降低,整合度和创新功能却逐步增强。生物世界最终形成的不仅是零件的叠加,而是一种多层次高度协同的有机整体。生命的合作网络之中,“个体”与“整体”往往难以绝对划界,但合作的广度和深度,是推动演化创新与生物多样性的核心动力。
从分子到生态系统
回到最初的问题:那片竹林算一个生物还是多个生物?其实,这并不是一个非黑即白的问题,而是取决于我们关注的层面和角度。
从遗传上看,地下茎相连的竹竿源于同一个受精卵,基因完全一致,可以看作一个克隆体;而从生理和功能来看,每根竹竿能独立进行光合作用和水分调节,彼此具有一定自主性;再从生态学角度,整片竹林又作为一个整体与环境相互作用,占据独特的生态位。
生物个体的界定,与研究目标紧密相关。研究遗传多样性时,我们可将竹林视为一个整体个体;研究功能与能量流动时,每根竹竿又可以看作相对独立的单元;观察生态系统动态时,则可把整片竹林看作一个生态“斑块”。
这样的多视角不仅适用于竹林,更适用于整个生命世界。从分子、细胞、器官,到个体、种群、生态系统乃至生物圈,生命在各个层次上都展现出不同程度的组织性与整合性。重要的不是在某一层级上寻找“真正的个体”,而是理解每一层级上协作与整合的机制与程度。例如,在单细胞层面,生命依赖分子机器完成自身维持;在多细胞层面,神经和内分泌系统协调各细胞;在物种间,共生现象促成跨物种的功能单元。这种层层嵌套、相互协作的结构,造就了生命体系的复杂和多样。
理解不同层面的生命整合,不仅加深我们对生命本质的理解,也推动了农业、医学和环境治理等实际应用——无论是利用根瘤菌减少化肥消耗,还是开发微生态医学新方案,抑或依托细菌生物膜处理污水,都是对生物多级整合理解的应用。
归根结底,生命世界的边界是模糊且动态的,各类生物组织只是整合程度不同的“有机体”。正是这种多层次、多样化的组织协作,使生命系统拥有无限的可能和惊人的创新力。真正值得赞叹的不只是物种的数量,更是生命在形态和功能上的多样组织方式,每一种层级的整合,都是演化智慧结出的硕果。