发育系统理论
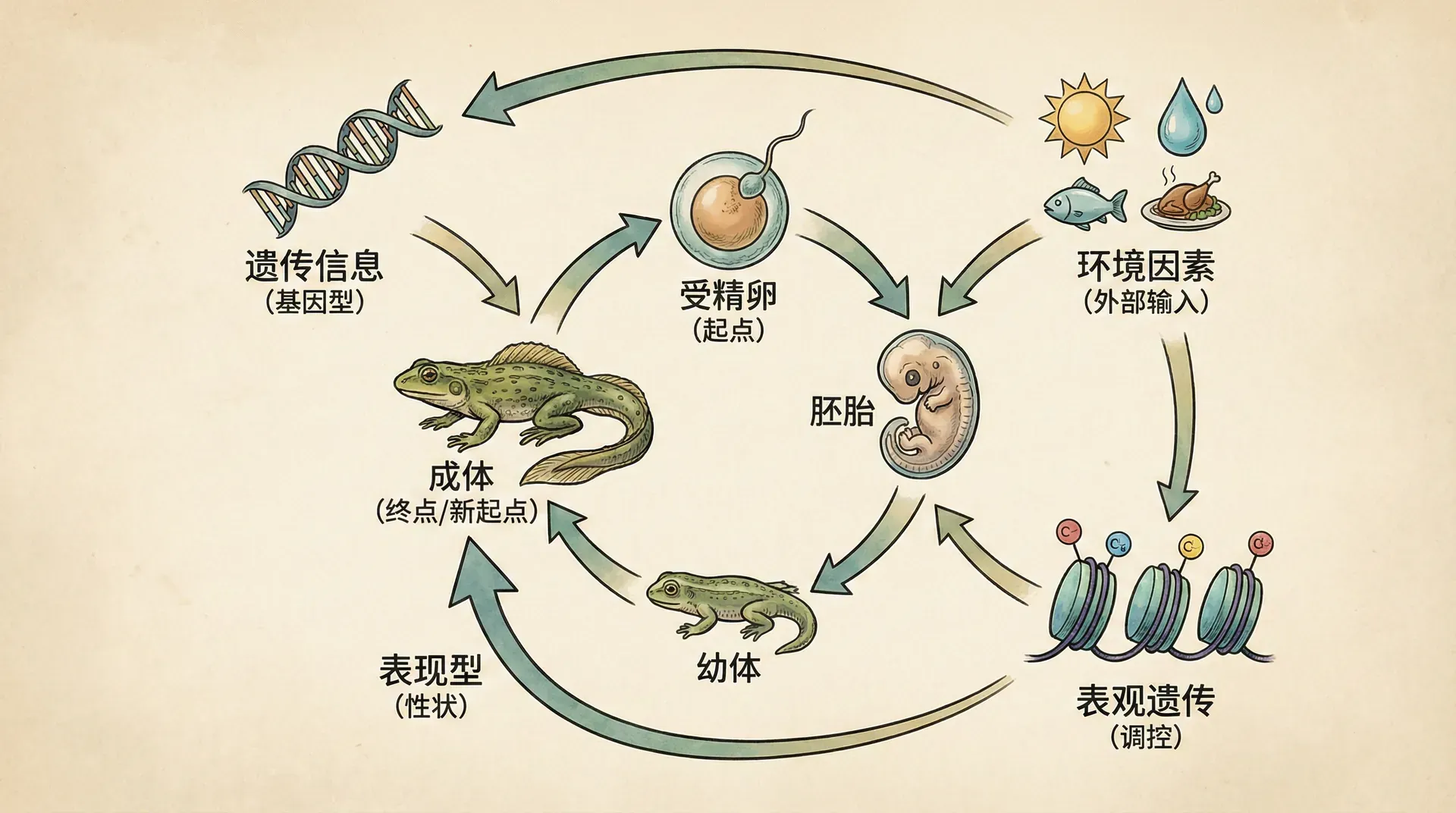
在生物学领域,一个古老的问题始终困扰着研究者:一个生物体的特征究竟是由基因决定的,还是由环境塑造的?这就是著名的“先天与后天”之争。然而,随着对生命发展过程认识的深入,科学家们逐渐意识到,这种非此即彼的思维方式本身可能就存在问题。发育系统理论(Developmental Systems Theory, DST)正是在这样的背景下应运而生,它为我们理解生命的发展提供了一个全新的框架。
发育系统理论的核心观点可以概括为:生物体的发育不是基因和环境的简单相加,而是一个动态的、持续展开的过程。在这个过程中,基因、表观遗传修饰、细胞环境、生态环境以及社会文化因素相互交织,共同塑造着每一个生命个体。这个理论强调,我们不应该把生物体看作一个由基因“编程”好的静态机器,而应该将其理解为一个不断与环境互动、在时间中展开的动态过程。
发育系统理论的关键洞见:生命的发育是一个过程而非产品,是基因组、表观基因组和发育环境共同作用的结果,任何单一因素都无法独立决定最终的表型。
从物体到过程
传统生物学常常将生物体看作一个静态“物体”,就如同根据基因蓝图装配而成的机器,但这种看法忽视了生命在时间维度上的连续变化。事实上,从受精卵的胚胎时期,到个体的出生、成长、成熟与衰老,生物体始终处于不断转变的过程中。发育系统理论主张,应当将整个生命周期视为分析生命的核心单元,而不是仅仅聚焦于某一固定时刻的状态。
以蝉为例,不同种类的蝉幼虫会在地下生活数年(有的可长达十七年),期间缓慢吸取植物根部的汁液。直到某一年夏天,具备足够的生存资源后,幼虫才会集体钻出地面,进行短暂的羽化和繁殖,然后生命周期结束。如果我们只看见地面上成群的成虫蝉声鼎沸,而忽略了漫长的地下阶段,就无法理解它们独特的生命周期和生存策略。正是多年地下蛰伏的过程,为短暂地面生活的繁衍爆发做了充足准备。
下表总结了蝉生命周期的几个关键阶段及其过程性特点:
这一例子生动展示了发育的过程性本质:个体当前的发展状态是之前各阶段积累的结果,同时也为未来的变化奠定基础。不同阶段间相互衔接,缺一不可,这种“过程依赖”意味着任何阶段的变化都将深刻影响生命周期的整体展现。
基因与环境的动态互动
长期以来,人们习惯于将基因和环境看作两个独立的因素,认为它们各自对生物体的发育贡献一定的“份额”。然而,发育系统理论指出,这种“基因+环境”的加法模型从根本上误解了发育的本质。真实的情况是:基因和环境始终处于动态的互动之中,它们的影响是相互依存、无法分割的。
美国心理学家丹尼尔·勒曼(Daniel Lehrman)在研究动物行为发育时提出了一个重要观点:真正与环境互动的不是基因,而是处于特定发育阶段的生物体本身。基因的表达效果取决于细胞环境,细胞的发育取决于组织环境,组织的形成又受到外部环境的影响。这是一个多层次、相互嵌套的互动系统。
研究发现,藏族人群携带EPAS1基因的特定变异,这个基因与血红蛋白的调控有关。但是,仅有这个基因变异并不能完全解释藏族人对高原的适应能力。实际上,藏族婴儿从出生开始就暴露在低氧环境中,这种环境刺激会激活一系列基因的表达,促进肺部血管的发育,增加血液中的氧气运输效率。如果一个携带同样基因变异的个体从小生活在平原地区,其身体不会发展出相同的生理特征。
这个例子展示了发育的“概率性表观遗传”特征。吉尔伯特·戈特利布(Gilbert Gottlieb)提出,发育的结果不是由基因预先确定的(预成论),而是在发育过程中逐步显现的(表观遗传论)。同样的基因型在不同的环境中可能产生不同的表型,因为环境条件影响着基因表达的时序和强度。
更重要的是,这种互动是双向的。大熊猫幼崽出生时极其弱小,体重只有母亲的千分之一左右,这在哺乳动物中极为罕见。这种“早产”状态意味着幼崽的许多器官系统需要在出生后继续发育。但这也给了环境更大的影响空间:母亲的哺育行为、巢穴的温度和湿度、以及幼崽与母亲的互动,都会深刻影响其神经系统和免疫系统的发育。研究发现,那些得到充分母婴互动的幼崽,其应激反应系统的发育更为健康,这种影响甚至可以持续到成年期。
在发育过程中,基因和环境不是各自独立起作用的“原因”,而是相互依存的“条件”。一个基因的效应取决于它所处的细胞环境和外部环境,而环境的影响也取决于生物体的遗传背景和当前的发育状态。
发育系统的三个组成部分
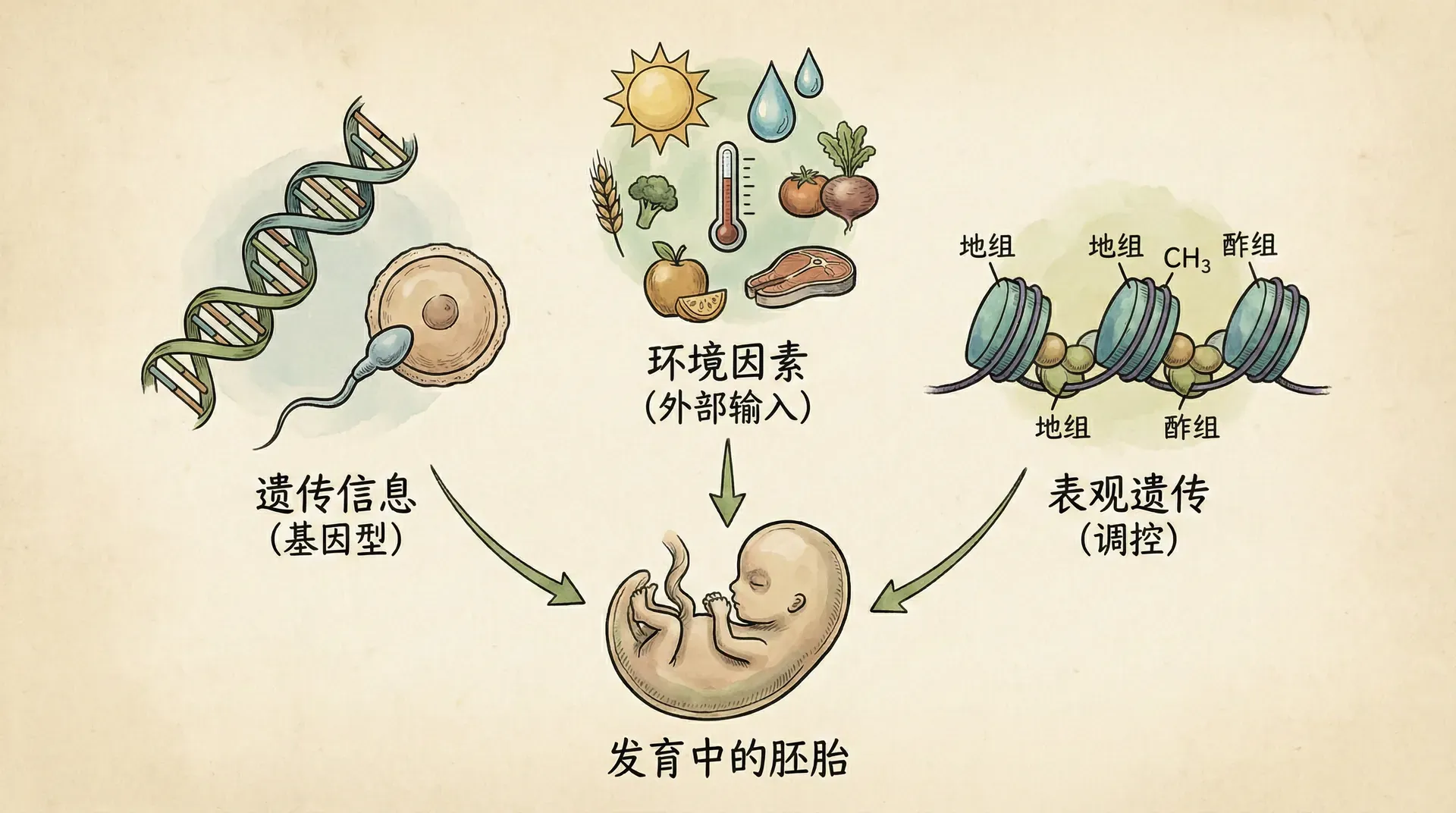
发育系统理论强调,生物体的生命周期能够持续、重建,依赖于一套由多个层面构成的“资源库”。这一理论提出:发育不仅仅依赖于基因本身,而是包括基因组、表观基因组和发育生态位这三个紧密关联、相互作用的系统,共同决定了生命的形成与延续。
让我们逐一梳理这三大组成部分:
-
基因组(Genome)
最常见的遗传信息载体,指的是DNA序列本身,包括编码蛋白质的基因以及各种非编码调控区段。它像是“零件库”和“初步设计图纸”,为生命活动提供基础。 -
表观基因组(Epigenome)
指附加在DNA及其相关蛋白上的化学修饰(如甲基化、组蛋白修饰)以及染色质的开放/紧密结构。这些修饰决定了哪些基因会被激活或沉默,并且可以响应环境信号动态变化。值得注意的是,一些表观遗传信息能够经由生殖细胞部分传递给下一代,成为遗传变异的新层面。 -
发育生态位(Developmental Niche)
由West和King等学者提出,指支持正常发育所需的外部环境资源与条件,包括物理环境(温湿度、光照)、营养、社会互动甚至文化传统。重要的是,这些资源很大程度上由父母或群体成员主动建构和维护,是个体发育中不可忽视的一环。
三种机制对比示意:
蜜蜂的分工机制:
- 女王蜂与工蜂都拥有基本相同的基因组;
- 幼虫期吃蜂王浆的个体通过表观基因组差异(甲基化等)发育为女王蜂;
- 蜜蜂的蜂巢结构和复杂社会行为(如舞蹈传递食源信息)构成了典型的发育生态位,为整个族群的持续繁衍提供“环境传递”。
发育系统理论让我们明白:遗传不仅仅是DNA蓝图的传承,还包括能被环境调控和部分传递的“调光系统”——表观遗传,以及可复制、可学习的发育环境本身。三者交互协作,保证了生命特征的可靠重现与灵活适应。
例如,现代水稻育种不仅重视品种的基因型,还会关注母本在形成种子时的田间环境(影响表观基因组),以及后续实际田块的土壤和水分管理(属于发育生态位)。在保护野生动物时,保护的不仅是基因多样性,更包括合适的栖息地和种群的社会知识。例如麋鹿的迁徙路线就包含了多代个体通过社会学习共同维护的发育环境。
表观遗传
以往的分子生物学将基因类比为“蓝图”,但现代表观遗传学告诉我们,发育更像是一场“现场演出”:基因提供演员名单,但是否上场、以什么方式上场要取决于环境中不断变化的指令。表观遗传是这套指令的分子基础。
表观遗传两大主流修改机制:
- DNA甲基化:让某些基因处于静默状态,通常位于启动子区域;
- 组蛋白修饰(如乙酰化或去甲基化):影响DNA包裹的紧密程度,决定基因的可读性。
这种修饰是可以动态变化的。例如,环境温度或营养状态就会影响特定基因的甲基化模式。
在水稻的表观遗传研究中发现,经历低温春化的水稻,关键开花基因启动子上甲基化水平升高,从而影响次年开花时间和产量,这也是农民长期“适地适育”的实践基础。
表观遗传“记忆”还可能跨越几代人。例如:
- 20世纪六十年代中国饥荒期间出生的人,在成年后罹患糖尿病、心血管疾病等的风险上升。研究显示其代谢相关基因的甲基化异常,这一表观标记部分传递给了下一代。
- 与此同时,在一些华北地区的沙冬青等植物,经历极端气候的植株会调整种子表观状态,使后代更耐旱。
表观遗传还是发育可塑性的分子桥梁。发育可塑性,指的是相同基因型在不同环境条件下能表现出不同的表型。例如:
- 蝌蚪变形: 黑斑蛙的蝌蚪在遇到捕食者气味时,更快变为腿发达、体型健壮的幼蛙。这一变化不仅涉及激素,还伴随着与新陈代谢有关基因的甲基化调整。
- 荠菜的分枝变化: 在土壤盐分升高时,荠菜发育出更多地上分枝,相关基因区域甲基化下降。
飞蝗的相变:
通过表观遗传调控,生物可以根据环境压力灵活“切换”发育路径,提高环境适应性。令人着迷的是,这种适应甚至可以为下一代“预设”更合适的起点。
表观遗传机制是DNA遗传与环境影响之间的桥梁。它让生物体“记住”环境线索,并通过化学修饰将这种记忆固定下来,有时甚至“写入”到下一代,使个体和种群更具灵活性与适应性。
发育的权衡与优化
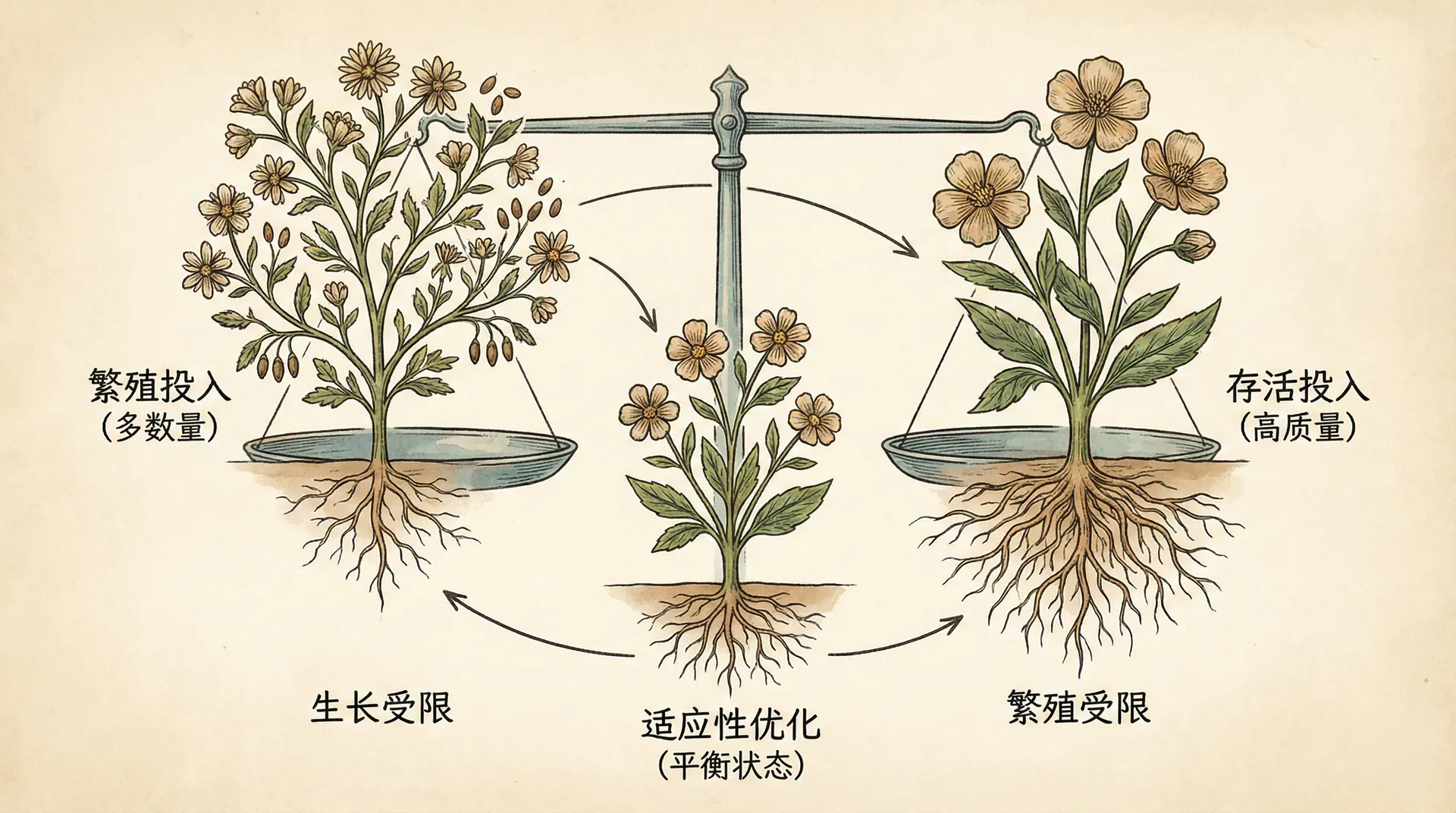
每一个生物,在其一生中都会面临最核心的难题:有限的能量和资源,究竟如何在生长、维持和繁殖之间进行最佳分配?生命史理论(Life History Theory)就是专门研究生物如何在不同环境压力下权衡这些分配策略的理论框架。从发育系统理论的角度来看,生物个体的生命周期就是一连串动态的发育决策,每一个关键节点都是不同目标(如存活、成长、繁衍、养育等)的平衡或取舍。
下面概括三大典型的权衡关系:
每个物种、个体都会根据自身遗传基础和生态环境,演绎出不同的生存与繁衍方案。我们来看几个具体案例,帮助理解这些权衡如何实际作用:
竹子的极端生命周期决策
毛竹等大型竹子是“一生只开花一次”型的典型代表。它们会长达几十年(60-120年)更倾向于投入资源发展营养体、扩展根状茎进行克隆繁殖,几乎不用于有性繁殖。只有在特殊环境信号下,才会“集体押宝”,同步开花并付出巨大代价:所有积累资源转而用于繁殖,完成使命后竹林整片死亡。这种极端取舍背后是对环境波动的应对:同步开花能集中产出大量种子,把存活的概率“分散到未来”。
北极狐与南美貂
权衡策略不仅体现在植物,也广泛存在于动物。例如,北极狐(Vulpes lagopus)和南美貂(Galictis cuja)都需要在食物稀缺季节抉择,是优先储存脂肪应对冬季,还是趁着夏季短暂的丰裕期争取多生几胎。北极狐往往更偏向于增加脂肪储备以保证存活,而热带的貂则侧重于加快繁殖节奏,因为生存危险主要在于捕食者而非环境恶劣。
蝉的“地下慢生长、地上快繁殖”模型
中国蝉类常年以幼虫形态蛰伏地下5-8年,摄入低营养的根汁,缓慢生长。在地下有庇护又安全,虽然成长速度慢,却大大降低了死亡风险。羽化为成虫后,仅存活数周至两月,需要极速完成交配和产卵。这就是“生长慢、繁殖快”的策略,事实上蝉羽化高峰每隔若干年就会出现,是对天敌“时间错位防御”的一种发育权衡。
除了极端案例,大多数物种的生命史策略更像是下表中的连续谱:
但现实中,大部分种群都介于这两极之间,并可表现出灵活调整。比如:
- 小型鸟类如麻雀,在繁殖季节昆虫丰富时会提升一次性产卵数,恶劣环境则减少数量,但提升亲鸟抚育程度。
- 鼹鼠在食物资源极端贫乏年份,会推迟甚至放弃当年繁殖,优先保存体力。
捕捞压力对鱼类发育策略的影响
鲫鱼的典型研究显示,在捕鱼压力大的水域,鲫鱼会提前性成熟、个体偏小。这是一种对死亡风险升高的快速响应。反之,在保护区,性成熟年龄推后、体型增加,提高了一次繁殖的后代产量。这里既有进化背景,也有发育可塑性:幼鱼感受到同伴“危险信号”时,将调整自己的发育轨迹。
无论是竹子的“押注开花”、蝉的“慢生长快繁殖”、还是鱼类对人类活动的快速变异,背后都体现了生物体对环境不确定性和资源有限性的自我优化。生命史策略并无绝对优劣,只有对具体情景的最优回答——而这正是发育系统理论强调的动态平衡与环境适应性。
生命史权衡揭示了一个重要事实:没有“完美”的生物。每一种策略都是特定限制下的最优方案,会带来某些优势,同时也意味着一些妥协。认清这些权衡,有助于深入理解生物多样性与保护策略的科学逻辑。
总结
发育系统理论为我们理解生命的发展提供了一个强大而灵活的框架。它超越了传统的"先天与后天"二元对立,将发育理解为基因组、表观基因组和发育生态位动态互动的过程。这个理论强调,生命不是基因的预设程序,而是在与环境的持续互动中不断展开的旅程。
从竹子的生长策略到藏族人的高原适应,从飞蝗的相型转变到蝉的生活史安排,我们看到了发育系统理论如何帮助我们理解生物世界的多样性和复杂性。更重要的是,这个理论为人类自身的发展、健康和福祉提供了新的视角,提醒我们关注生命早期的经历、重视环境条件的作用、理解发育的可塑性,并在各种实践领域做出更加科学和人道的决策。
生命是过程,不是物体;是可能性,不是宿命;是系统,不是部件的简单集合。这就是发育系统理论带给我们的核心洞见,也是我们在面对21世纪的生物学、医学和环境挑战时需要铭记的智慧。