生物个体性与共生关系
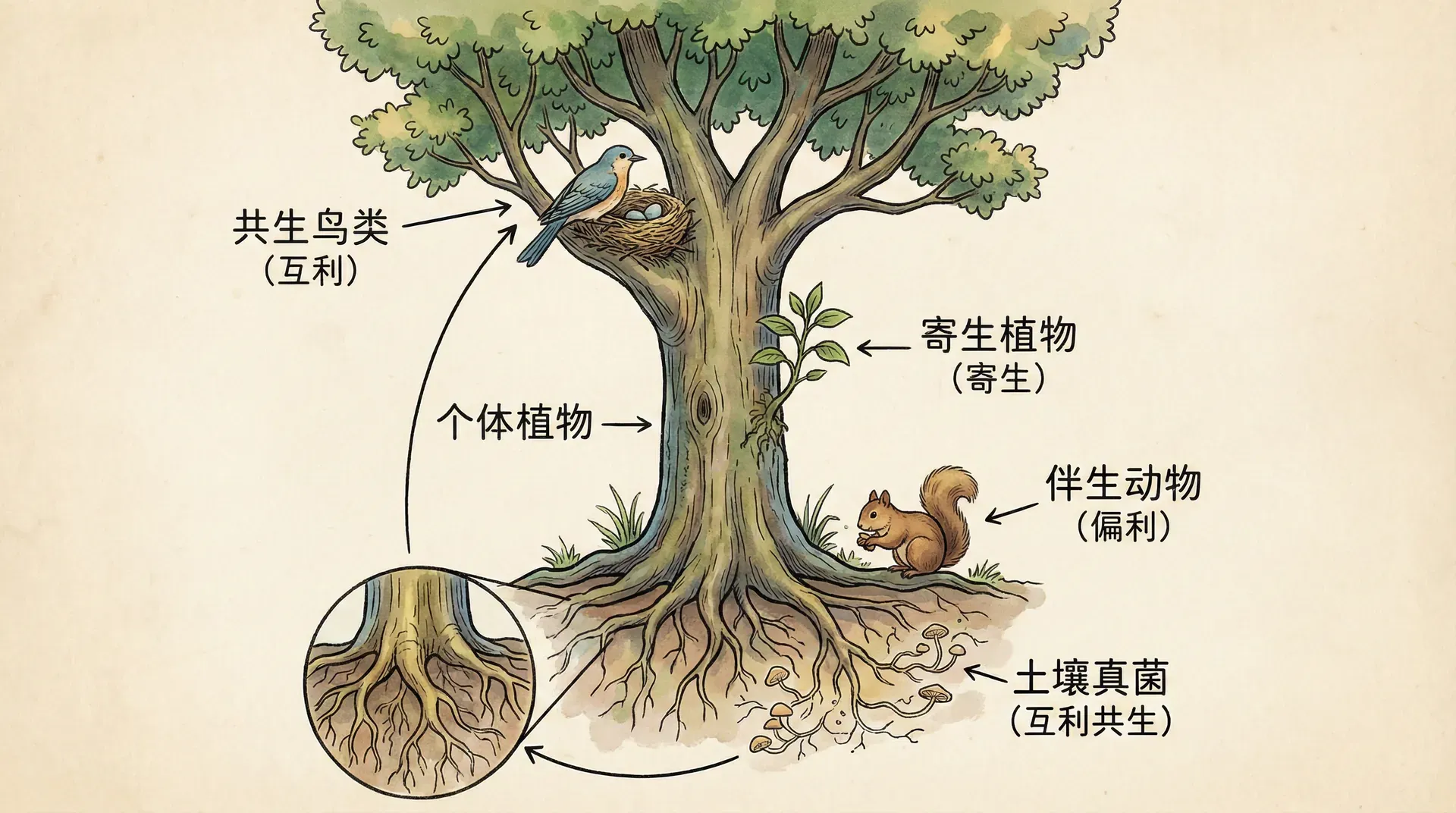
在生物学的学习过程中,我们往往认为“个体”这个概念是不言自明的。一只蚂蚁是一个个体,一棵树是一个个体,一个人也是一个个体。然而当我们深入观察自然界时,就会发现这个看似简单的问题其实充满了复杂性。
当你观察一个蚂蚁巢穴时,你看到的究竟是什么?是数千只独立的蚂蚁个体,还是一个由众多成员组成的“超级有机体”?工蚁、兵蚁、蚁后各司其职,它们的分工如同我们身体中的不同器官。如果蚁群可以被视为一个整体,那么判断“个体”的标准是什么?
这个问题在现代生物学研究中变得越来越重要,特别是当我们发现许多生物的生存和功能依赖于与其他物种的紧密合作时。在田间地头,大豆植物依靠根部的细菌来获取氮素;在南方的朽木中,白蚁需要肠道里的原生动物才能消化纤维素;在深海中,乌贼携带着发光细菌来躲避捕食者。这些生物究竟应该被看作独立的个体,还是作为一个功能单元的整体?
自然选择与适应的基本原理
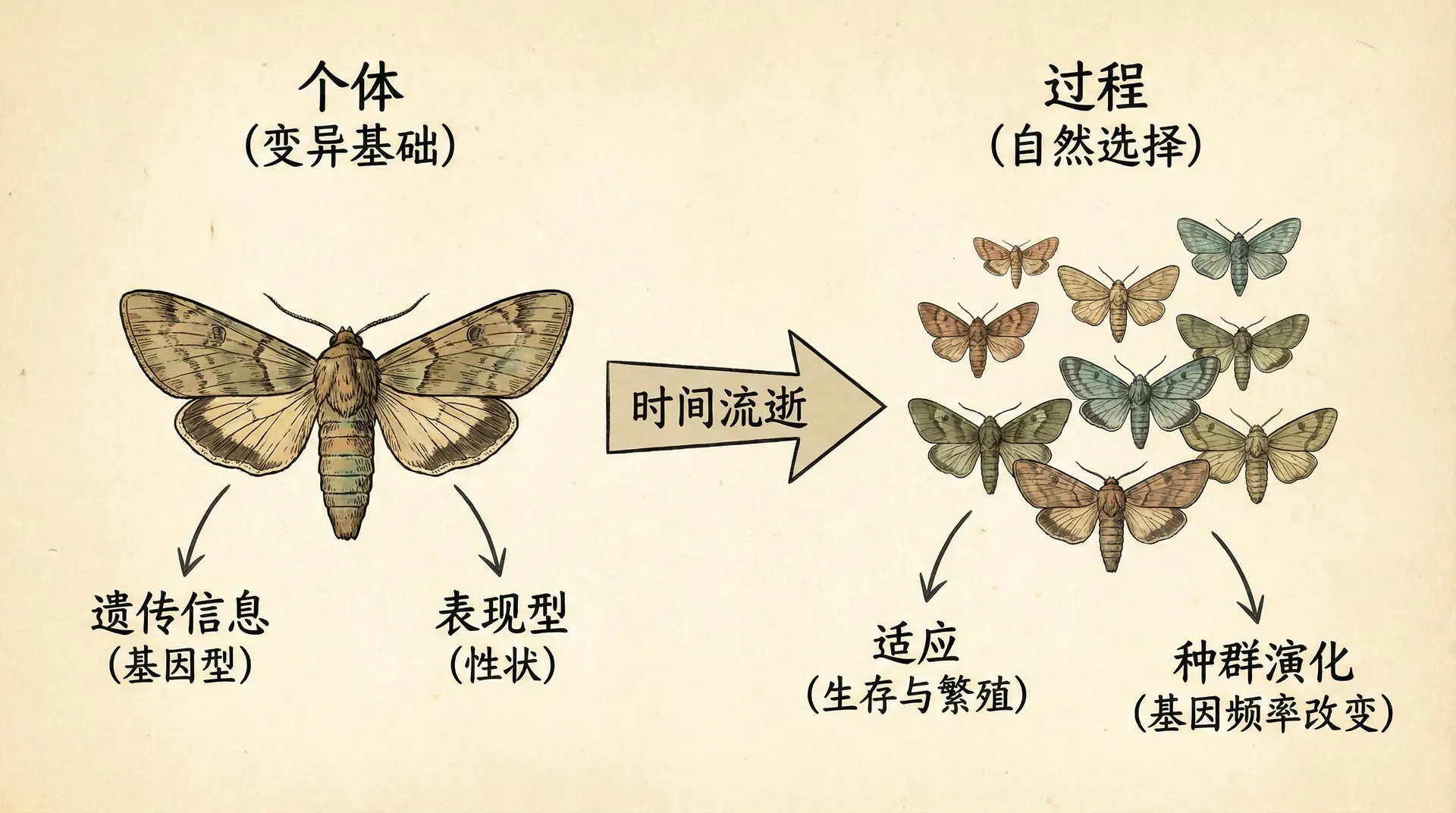
要理解生物个体性问题,我们需要从自然选择和适应的基本机制入手。达尔文进化论认为,有利的遗传性状会在环境选择压力下积累,从而形成我们今天所见的“适应”现象。
以下方图表为例,假设在华北地区野兔种群中,既有浅色毛的个体,也有深色毛的个体。冬季雪地环境下,浅色毛的兔子更能与环境融为一体,减少被捕食的风险,从而其存活率更高。图中“浅色毛野兔数量”随世代增加不断上升,“深色毛野兔数量”则逐代下降。这正体现了具有优势性状(浅色毛)的个体通过自然选择在种群中逐渐增加。
在进化生物学中,这个过程可以拆解如下:基因(复制子)存在变异,不同的个体(互作子)因基因差异表现出不同的毛色。浅色毛的兔子生存和繁殖更成功,其相应基因在下一代中的比例也随之升高。世代更迭后,浅色毛的基因频率不断累积,种群特征逐渐改变。
正如图表数据所示:世代更迭下,能更好适应雪地环境(浅色)的兔子大幅增加,而不适应的(深色)逐渐被淘汰。这体现了自然选择“筛选”基因和表型的过程。
自然选择的核心体现在个体间存活与繁殖成功率的差异。最终,优势性状的基因更易在种群中累积,表型分布随之改变。值得注意的是,这里的例子假设基因型和表型——以及个体与基因的关系——是一一对应的。因此,浅色毛这个性状与相应基因、个体及其后代之间有直接关系。但在多物种合作系统中,这一对应关系经常会变得复杂甚至模糊,这也是我们后文将要探讨的核心难题。
共生关系带来的挑战
当一种生物体的适应性特征并非完全源自自己的基因,而是与其他物种的合作成果时,传统进化理论的“个体”与“适应性”范畴就变得暧昧。中国的自然和农业实践中,这类现象屡见不鲜,挑战了我们对生物个体性的常规理解。
大豆与根瘤菌
中国东北广阔的大豆田是这个故事的发生地。每年春天,农民们种下大豆,却并不担心土地是否肥沃。秘诀就在于大豆根部长出的根瘤—— 那些小疙瘩,其内部住着一类神奇的细菌:根瘤菌(Rhizobium)。
- 双方分工:大豆通过根瘤给予根瘤菌温暖、低氧的家园和通过光合作用获得的糖分。根瘤菌则把空气中的氮气转化为大豆能吸收的含氮营养,这一过程叫“生物固氮”。
- 传递方式:大豆种子本身不携带根瘤菌。每一代新大豆都要从土壤环境中重新“招聘”这些细菌“员工”。
- 归属之谜:因此,所谓“固氮能力”其实不是大豆自己的基因直接带来的,而是跨物种合作的产物。它究竟属于谁?
白蚁与肠道共生微生物
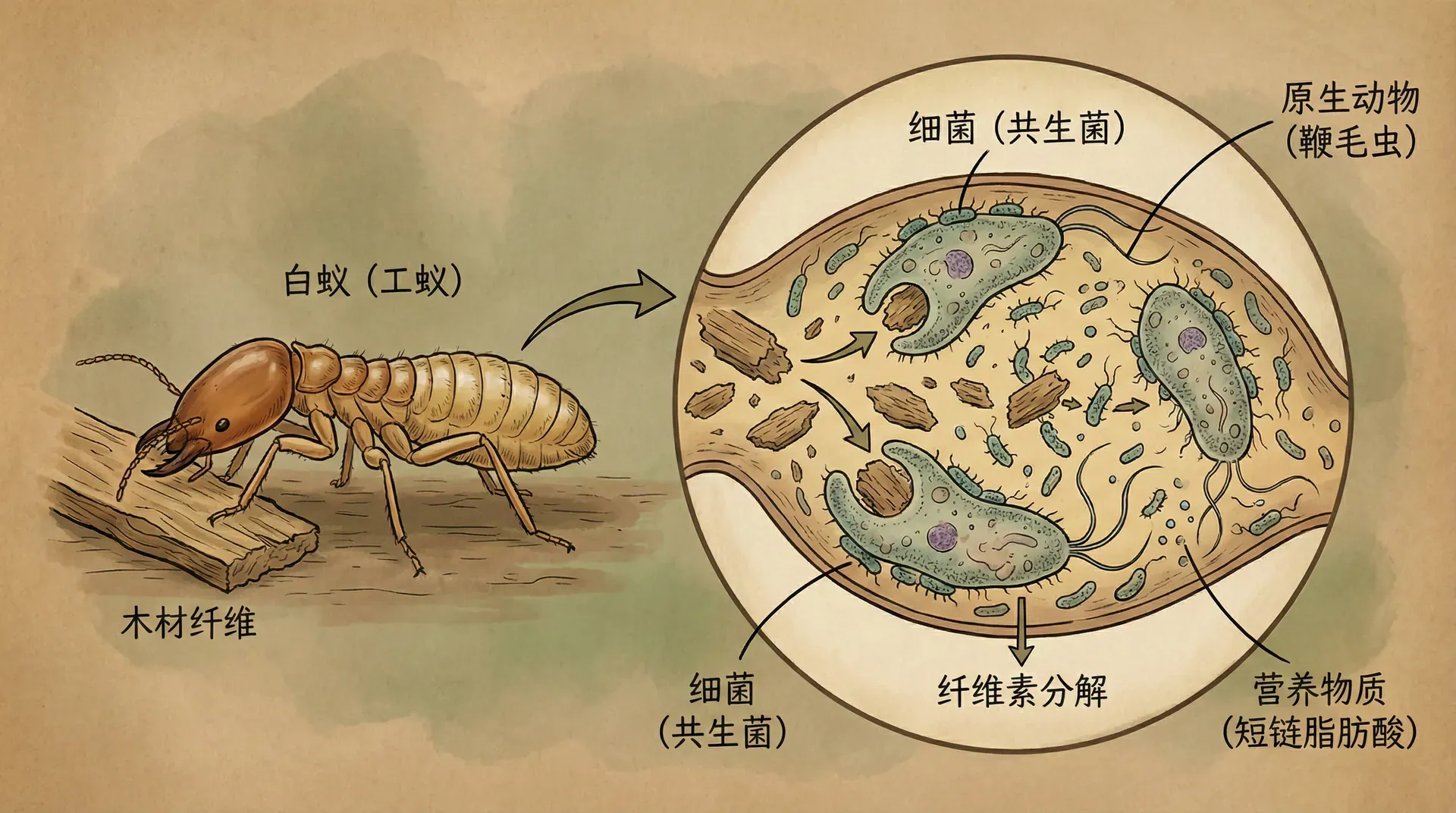
南方省区的土壤和枯木中,成群的黑翅土白蚁(Odontotermes formosanus)靠啃食木头为生。但木头的主要成分是结构坚固的纤维素,绝大多数动物无法自行降解。白蚁的“秘密武器”来自肠道中的共生原生动物、细菌和真菌。
- 合作机制:原生动物(如鞭毛虫)能分泌纤维素酶,将纤维素分解。共生菌进一步协作,产出白蚁可以吸收的有机分子。
- 获得方式:白蚁幼虫孵化时肠道是无菌的,必须通过与成年蚁之间的“肛门喂食”行为获得完整的微生物群。否则,它们即使有食物入口,也因无法消化而死。
- 个体性迷思:离开肠道群落,白蚁本体其实无法获得能量。消化这个生物特征,是整个“白蚁-微生物体”组合体的属性。
常见消化共生关系一览表:
红树林螃蟹与硫细菌
以红树林生态系统为例,红树林拳击蟹(Perisesarma bidens,分布于中国南方沿海)能在低氧、高硫的泥滩中生存,这依赖于其鳃腔内的硫细菌。这些细菌能氧化硫化物,为螃蟹提供部分能量,有助于它们在极端环境下生存。
- 互助内容:螃蟹呼吸时吸入水中硫化物,细菌将有毒的硫化物转化,并释放能量和较为无害的产物。
- 共生方式:细菌长期定植于螃蟹鳃腔,但在严重污染或环境突变后会显著减少,有时需要重新获得。
- 功能整合度:功能更多限制于螃蟹鳃部,个体边界相对保持,但一旦丧失细菌,适应能力骤降。
共生系统对比结构
共生现象说明,适应性特征常常依赖于多物种密切合作。个体与基因的“一一”对应关系在这些场景中失效,进化解释必须承认功能和遗传的多重交错。
在这些共生系统里,一个看似“个体”的实体,其适应特征可能需要跨物种的遗传合作与每代循环协作,也许这正是“生物个体”需要重新界定的根本原因。
重新定义生物个体
面对共生带来的混乱,生物学家提出了新的概念框架。Wilson和Sober(以及许多后来学者)给出了如下定义:
生物个体指的是在功能整合(functional integration)和共同命运(shared fate)两个维度上高度耦合的生命系统。
功能整合
功能整合指系统内部各组成部分间的协同配合与相互依存。它既可以通过器官、细胞结构的分工体现,也可以通过行为与代谢的高度协作展现。
各器官密不可分:
功能整合呈现连续梯度:从零协作(如沙漠里独立仙人掌),到松散互动(蜜蜂与花),再到协作紧密、不可分割的有机体(人、白蚁-微生物联合体)。
共同命运
共同命运指系统组成部分的生死兴衰是否被紧紧绑在一起。如果一个部分的灭绝通常伴随整个系统功能的终止,那么这个系统命运的“粘合度”就极高。
举例列表:
高度共同命运
- 人体内的心脏与肝脏,个体生死与功能紧密捆绑;
- 白蚁与肠道微生物,离开共生体,彼此都难以独立续命。
中等共同命运
- 大豆与根瘤菌,部分绑定,根瘤菌可在土壤独活,但大豆遇到贫瘠则依赖根瘤;
- 拳击蟹与硫细菌,环境变化时一个可暂时失去另一个但影响适应力。
低共同命运
- 森林中不同树木,各自独立生老病死。
个体性的连续谱
个体性既包括功能整合,也包括共同命运。两者的不同组合,塑造出一系列从松散群落到高度一体化系统的过渡形态。
- 极端一体化:“兔子”“人”等多细胞动物,各系统命运与功能均牢牢绑定。
- 中间类型:各种形式的共生联合体,整合和命运都有不同程度的耦合。
- 极端松散:如过路的蚁群、森林植物垄断个体,彼此结合极低。
认识到个体性不是简单的“Yes/No”,而是一个多层次、多维度的连续梯度,有助于我们揭示生命多样性、理解进化过程中的“混合体”以及古老共生关系如何最终转化为统一的有机体。
这种理解对现代进化生物学极为重要。不少今天看似“一体化”的有机体——比如真核细胞中的线粒体、叶绿体,起源其实都可以溯源为古老共生。当各种共生体协作临界到足够紧密时,个体性边界会发生活跃的重构。
事实上,我们自己就是最典型的“多古老生物整合体”——无数的细胞器、基因模块,跨越亿万年的共生与整合,才形成今天的“人类”这个多细胞个体。今天的共生,或许就是未来“新个体”的萌芽。
瞬时性与间歇性的个体

南海发光乌贼的例子引出了生物个体性的另一个复杂维度:时间性。不是所有的生物个体都是持续存在的,有些个体会在时间轴上反复出现和消失。
发光共生体的日周期
让我们仔细追踪一只发光乌贼在一天中的状态变化。
清晨时分,太阳升起,乌贼结束了一夜的觅食,沉入海底休息。此时,它的发光器官中仍然有大量发光细菌,但随着乌贼收缩特殊的肌肉组织,大部分细菌被排出体外,只有约5%留存在发光器内部的隐窝中。细菌密度骤降,发光停止。
白天,乌贼休息,体内残留的细菌开始繁殖。乌贼为它们提供营养丰富的培养液,其中含有糖类和氨基酸。细菌数量逐步增长,从大约10万个细胞增长到接近1000万个。
黄昏时分,当细菌密度达到临界点——每毫升约10⁷个细胞时——群体量感应机制启动。细菌释放一种叫做“自诱导物”的信号分子,当这种分子的浓度达到阈值,所有细菌几乎同时激活发光基因。发光器开始发出淡蓝色的光。
夜晚,乌贼游动觅食,发光器持续工作。通过调节发光器开口和内部反射结构,乌贼能够控制光线的强度和方向,使其与月光和星光相匹配,消除自己的影子。这个精确的伪装使乌贼能够安全地在开阔水域活动。
第二天清晨,循环再次开始。
这个图表展示了24小时周期内,发光细菌密度和发光强度的变化。细菌密度在白天逐渐上升,而发光只在密度达到阈值后才开启,呈现出明显的间歇性。
个体的“闪烁”
这个日周期模式带来了一个深刻的问题:我们应该如何理解这个共生系统的个体性?
一种观点是,每天都有一个新的“乌贼-细菌”共生个体诞生。清晨排出细菌后,旧的共生个体消失;傍晚发光开启时,新的共生个体形成。按照这个观点,一只乌贼在其生命周期中会与数百个不同的细菌群体形成数百个不同的共生个体。
另一种观点是,有一个持续存在的共生个体,其功能整合程度在一天中有规律地波动。就像我们睡眠时意识活动降低,但我们仍然是同一个人一样,发光功能的关闭并不意味着共生个体的消失,只是整合程度的暂时降低。
从生物学功能的角度看,第二种观点似乎更合理。虽然细菌群体的具体成员在变化,但有几个关键事实支持个体的连续性。首先,每天留存的“种子”细菌维持了遗传的连续性。其次,发光器官作为培养细菌的特化结构是持续存在的。第三,整个系统的调控机制——包括营养供应、密度控制、光学结构——作为一个整体是连续的。
我们可以把这类共生个体称为“间歇性个体”或“闪烁个体”。它们的个体性不是恒定的,而是在时间轴上波动的。这类似于灯泡的闪烁,虽然光亮时有时无,但灯泡本身持续存在。
时间尺度的错配
间歇性个体还揭示了共生系统中一个更深层的复杂性:不同组成部分运行在不同的时间尺度上。乌贼个体的寿命约为一到两年。在这段时间里,它经历了几百个发光周期。
单个发光细菌的寿命只有几个小时到一天。在乌贼的一生中,其发光器内经历了成百上千个细菌世代的更替。而发光器官作为一个结构,从乌贼幼年定植细菌时开始发育,一直持续到乌贼死亡。
从基因的角度看,时间尺度的错配更加明显。乌贼的基因通过乌贼的繁殖传递,世代周期以年计算。细菌的基因通过细菌的繁殖传递,世代周期以小时计算。然而,“发光伪装”这个适应性特征是两套基因系统共同作用的结果。
在间歇性个体中,不同组成部分在不同的时间尺度上变化。这意味着“个体”的边界不仅在空间上模糊,在时间上也是动态的。
这种时间上的复杂性在其他共生关系中也存在,只是程度不同。大豆植株存活一个生长季,而根瘤中的根瘤菌可能经历了几十个世代。白蚁工蚁存活数月到数年,而其肠道中的原生动物也在不断繁殖和死亡。
这些观察提示我们,在思考生物个体时,不能只考虑空间上的边界(哪些部分属于同一个体),还要考虑时间上的连续性(个体如何在时间中持续和变化)。对于共生系统,个体性既是空间整合的产物,也是时间过程的产物。
进化理论中的个体与过程
通过对共生关系和间歇性个体的讨论,我们逐步认识到“个体性”在生命系统中远比直觉所想更加多样和复杂。边界模糊、定义灵活,但这种复杂性恰恰促使我们反思:“个体”这个概念是否仍然有价值?我们能否仅通过“生物过程”来解释生命现象?
个体与过程
从某种角度看,每一个“个体”其实就是众多生物过程的一个临时交汇。例如:
这些实例表明,个体经常是多种过程的“节点”,表面看起来是稳定的单元,实则不断接受和参与各类生理、生态、进化过程。如果我们一味转向过程视角,似乎能更动态地把握生命的本质。
但现实更为细致。在进化生物学的语境下,个体仍然是核心概念。这是因为自然选择作为生物多样性和适应性产生的机制,其运作基础恰恰依赖于“个体”这一选择单元。
自然选择依赖于个体
自然选择发生有三个基本要素:
- 可区分的单元 —— 要有明确的生物实体,可以各自识别;
- 性状差异 —— 各个体表现出可遗传的差异;
- 不同的生存、繁殖结果 —— 差异导致适者生存、劣者被淘汰。
我们可以梳理自然选择的具体单元与过程的关系:
可见,在具体的自然选择场景下,我们始终需要有一个可以“比较、复制、淘汰”的载体。这个载体——个体,可以是单一生物体,也可以是高度整合的多物种复合体,但其“可识别性”和“竞争性”是基础。
过程视角的局限与个体的不可或缺性
如果完全放弃“个体”,只用“过程”解释生物进化,会遇到两个难题:
- 选择对象消失:过程可以描述现象如何发生,却难以明确描述“谁”在被选择。
- 适应丧失主体:适应性特征本身依附于具体生物个体,没有承载体,“适应”这一概念将无处附着。
例如,当我们说“发光是乌贼(及其细菌)的适应战略”时,背后隐含着存在一个具体的生物单元(不论多样性如何),正是它承载/表现了这一特征。如果仅仅用过程描述,无法说明“适应”归属于哪个实体。
个体与过程的有机结合
因此,个体与过程实际上是生命系统中不可分割的两个维度:
这种关系可以理解为:“个体是在过程中的暂时产物,同时又是过程得以实施的载体。”个体的边界常常因过程的变化而移动,但自然选择发生时,仍然需要界定出可以淘汰或复制的单元。
生物个体并非与过程无关的静态存在,而是在持续进化过程中被构建和维护的动态系统。理解个体性,必须同时关注它的结构特征与生成、维持它的各类生物过程。
对于共生系统尤其如此。我们既要识别作为选择单位的个体——无论是单一物种的生物体,还是高度整合的多种生物复合体;又要追溯和分析塑造这些个体的多尺度多阶段进化过程。只有将两者结合,才能充分解析共生体的适应性如何形成与维持,并为生命多样性的理解提供坚实理论基础。
总结
生物个体性是一个基础但复杂的问题。中国本土的共生案例——如大豆固氮、白蚁消化协作、南海发光共生——展现了个体边界的多样性和灵活性。对于多物种共生系统,传统的“一个生物体一个个体”的看法已无法解释复杂实际。适应性可能来自多物种的基因协同,功能整合和共同命运重新定义了“个体”。个体性有高有低,是一个连续谱,随功能协调和命运依赖而变化。
一些共生体的个体性随时间波动,呈现出“闪烁”特性。系统不同组成部分还可能运行在不同的时间尺度,使判断更复杂。尽管如此,个体概念在进化生物学仍不可或缺。自然选择和适应都需要具体的可区分单元作为载体,因此理解生命要兼顾个体和过程。
深入认识个体性也有现实意义:农业上可优化固氮利用,生态保护中能更清晰识别多样性单元,医学上则加深对“人体-微生物组”等复合系统的理解。
总之,共生关系显示生命的组织层次极其丰富,各层次间存在大量中间状态。承认和研究这些复杂性,是现代生物学的核心任务之一。