生物过程的动态特性
当我们观察一只蚕宝宝从孵化到吐丝结茧,再到最终破茧成蝶时,很难相信这些截然不同的生命形态竟然属于同一个生物个体。从体长仅有几毫米、整天啃食桑叶的毛毛虫,到无法进食、只能依靠体内储存营养的蛹,再到长有翅膀、专注于繁殖的成虫——这种变化之剧烈,远超我们日常生活中对“变化”的理解。
这种现象在生物界并非个例。长江流域的中华鲟从受精卵发育到成鱼需要经历十几年,期间体重增长超过百万倍;四川的大熊猫幼崽出生时体重仅为母亲的千分之一,粉红色、无毛、双眼紧闭,与我们熟悉的黑白相间的成年大熊猫判若两物。这些现象都指向一个生物学的基本事实:
生命是一个持续变化的动态过程,而非静止不变的状态。
理解生物系统的这种动态特性,对于我们把握生命现象的本质至关重要。传统观念往往将生物体视为具有某些固定特征的实体,但现代生物学研究越来越清楚地表明,从分子水平的代谢循环,到个体发育的形态转变,再到种群演化的长期变迁,变化与动态平衡才是生命的常态。
生物系统的多层次动态性
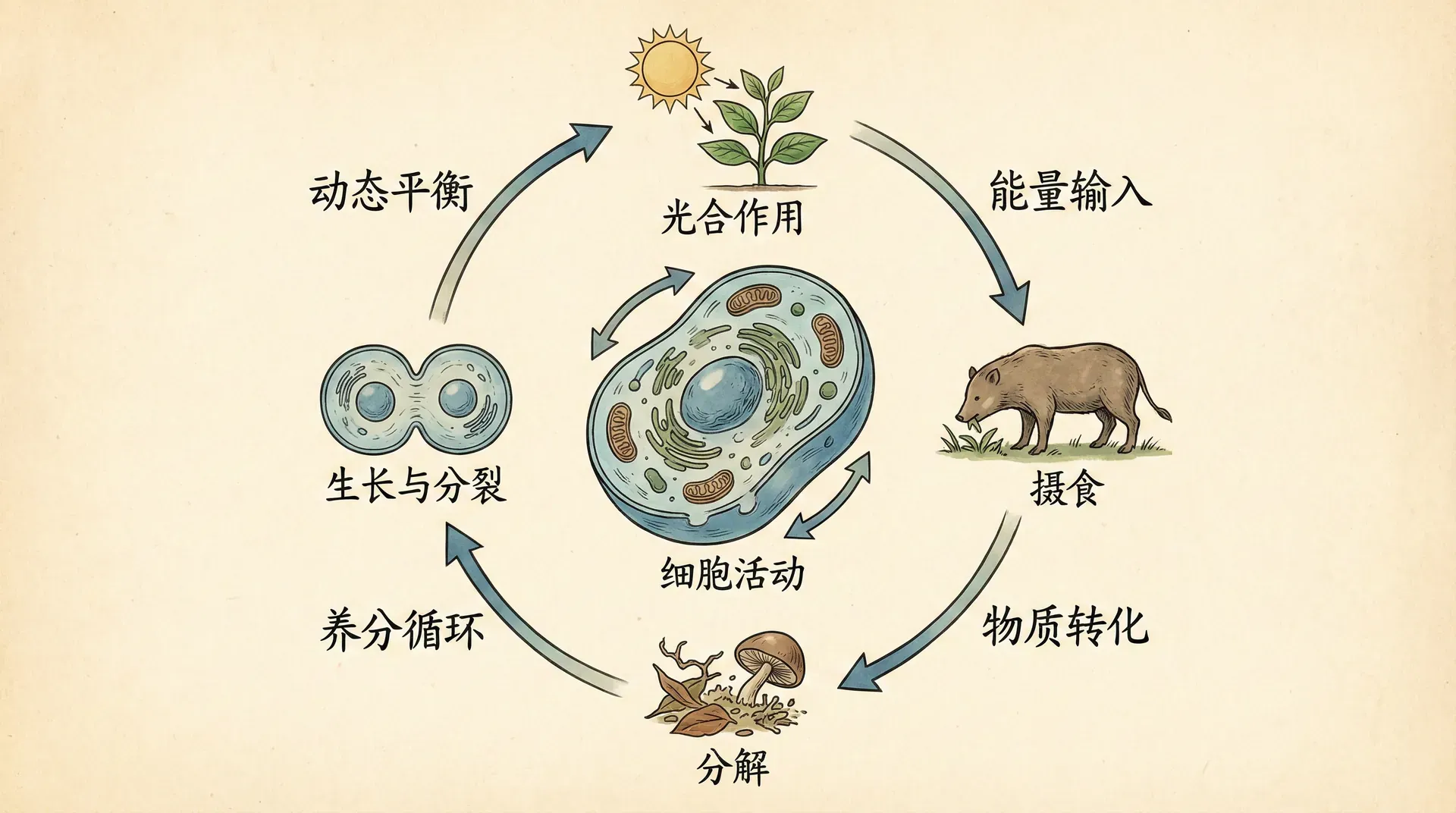
生命的动态特性体现在分子、细胞、个体乃至种群多个层次,并展现于从极短到极长的各种时间尺度上。以细胞为例,细胞内的生化反应网络复杂且快速:数以千计的代谢反应每秒钟不断发生。酶活性的高低会受底物浓度、温度和pH的微小波动影响,ATP分子的合成与消耗在细胞线粒体内循环不息,蛋白质持续被合成并降解,这些变化共同维持着细胞的动态稳态。
下方信息概括了分子水平上一些常见的动态过程:
在个体发育的时间尺度上,生物从受精卵到成熟个体,形态与生理结构发生巨大改变。例如,青蛙的生活史极具代表性:蝌蚪时期生活在水中,以藻类为食,依靠鳃呼吸和尾巴游动;进入变态发育后,蝌蚪逐渐长出四肢,尾巴缩短直至消失,肺部发育完善,转向陆地生活,消化系统也发生剧烈变化以适应肉食习性。类似的变化也出现在蝴蝶、蚊子等完全变态昆虫的整个生命周期中,这些过程在本章下文有更详细分析。
除了个体发育,生物的动态还体现在更长的演化时间尺度上。动物和植物的物种并不是恒定不变的,而是遗传变异和自然选择作用下不断演化的谱系。比如,现今中国的金丝猴与百万年前的祖先已经在形态、代谢和社会行为等方面出现了明显分化,而其中的多样性和变异性为持续进化提供了“原材料”。
不同时间尺度和层次的动态变化通常不是孤立存在的,而是互相耦合、彼此影响。例如,环境温度的随机波动可能直接影响细胞酶的活性(分子层次),进而影响个体的代谢效率与发育节律(个体层次),最终影响该物种在环境中的生存和繁殖表现(种群层次)。
这种多层次、多时间尺度的动态性对我们理解生命现象提出了理论和实践上的挑战。下方信息从蚕卵到成虫,外观、细胞甚至基因表达都发生了剧变,但我们依然视其为同一个体。这种“变与不变”的悖论是理解生命现象的核心问题。
发育过程中的形态转变
生物发育中最引人注目的现象之一,就是某些物种在生命历程中经历的巨大形态转变。这种转变的程度之大,以至于如果不了解它们的生活史,我们很可能会将同一生物的不同发育阶段误认为不同的物种。
昆虫的完全变态
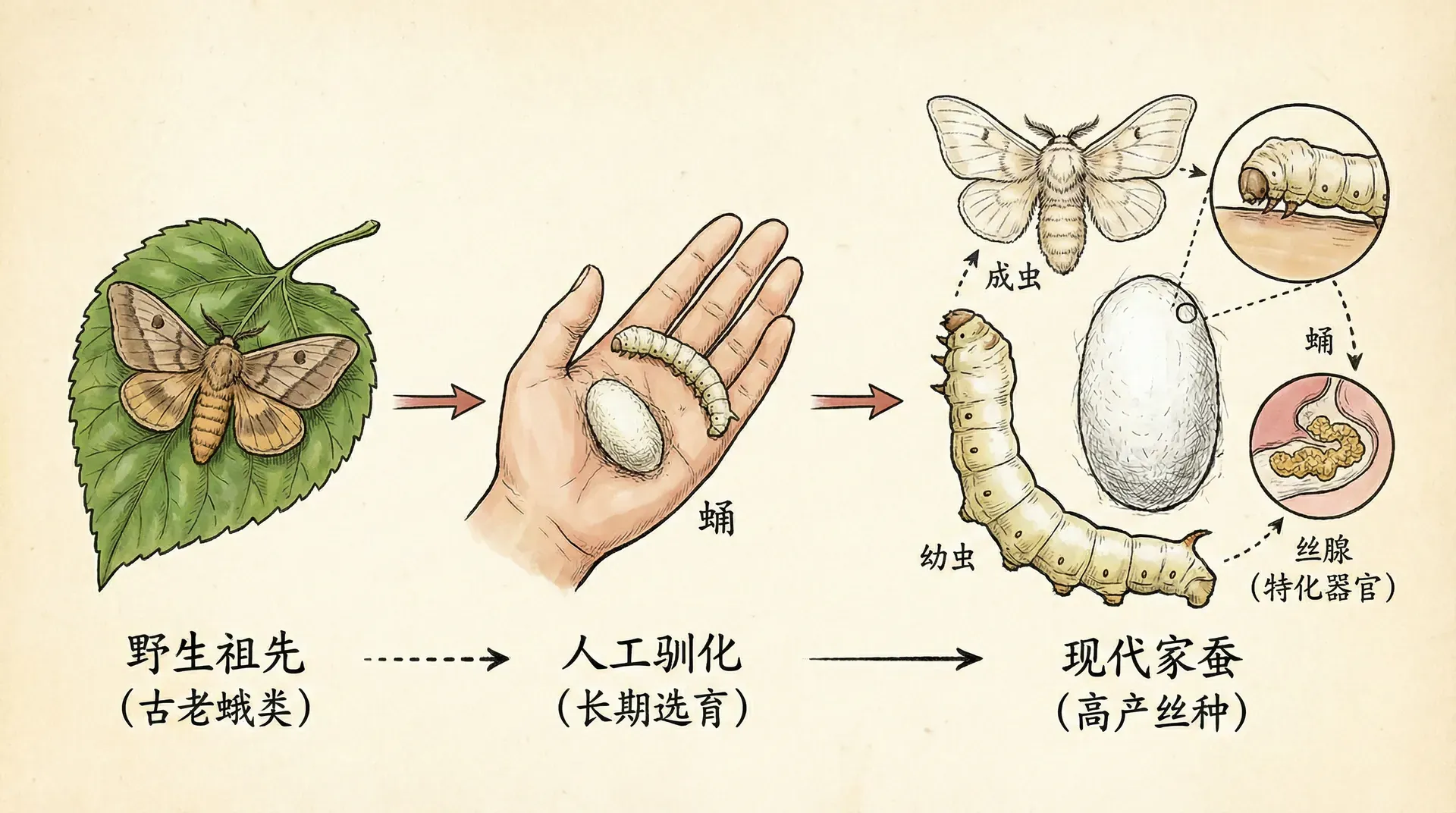
家蚕是中国传统养殖的重要经济昆虫,也是研究完全变态发育的经典案例。从春季孵化开始,一只家蚕要经历卵、幼虫、蛹、成虫四个截然不同的生命阶段。
家蚕幼虫在孵化初期体长仅约2-3毫米,体重极轻,刚出生阶段的体重约0.01克。随着天数的增加,体重的增长呈现出明显的阶段性:前5天增长缓慢,第5天前后约为0.25克;第5天至第15天为快速生长期,体重迅速上升,第15天左右可达4.25克左右;第15天至第25天增速有所减缓,体重增长至6克左右;25天以后,体重增长变得极为缓慢,逐渐进入成熟准备阶段。这一增长模式具有明显的“指数式”特征。
从上面的图表可以看到家蚕体重的变化过程:孵化初期为“慢速增长期”,随着发育进入第5天—第15天后,体重进入“爆发式快速增长阶段”,到20天左右已远远高于早期,体重差异悬殊。第25天后体重增幅减缓,接近熟蚕时达到6克左右,可比孵化初期增长近万倍。此时蚕幼虫停止进食,准备吐丝结茧。
化茧为蛹后,家蚕进入“形态重建期”:表观上看不见体重显著变化,实际上内部结构正在发生剧烈重组,绝大多数幼虫时期的组织被分解,成虫的结构逐步构建完成。这一阶段历时约12-15天。
最终羽化出来的家蚕成虫比幼虫时期体型变化显著:体长约2-3厘米,有两对带鳞翅膀,但因长期人工选育已几乎丧失飞行能力。成虫口器退化,无法进食,全部能量都来自幼虫时期积累的储备。成虫阶段生命极为短暂,仅3-5天,唯一任务是交配与产卵,随后死亡。
整体来看,家蚕体重的剧烈变化与发育各阶段密切对应,充分体现了生命现象的动态特性。
根据上方的体重变化图表,家蚕的各发育阶段在持续时间、体长及体重等方面具有明显差异,且与体重增长速率高度相关。下表对照该图表所示体重变化,对各阶段主要特征进行了归纳总结:
结合图表,可以清晰看到家蚕体重在0-5天“慢速增长期”之后,进入5-20天的“爆发式快速增长期”,25天后体重增速明显减缓,准备化蛹。蛹期虽然外观变化不大,但内部组织进行巨大重建。成虫阶段则体重减少,专注繁殖,生命周期完成。
这种发育节律和体重变化密切相关,是完全变态昆虫(包括家蚕、蝴蝶、蜜蜂等)的显著特征。其生态意义在于,幼虫阶段专注能量积累与生长,而成虫阶段专注繁殖和扩散,实现了同一物种不同发育阶段资源和环境的分化利用。
寄生虫的复杂生命周期
完全变态昆虫的发育已经显示出生物个体巨大而剧烈的变化,但某些寄生虫的生命周期则更为复杂。以华支睾吸虫(俗称中华肝吸虫)为例,这是一种在我国南方淡水鱼产区常见的肝脏寄生虫,其生命周期涵盖三类不同的宿主,跨越多个形态阶段。
整个生命周期可以按照“人(终宿主)→淡水螺(第一中间宿主)→淡水鱼(第二中间宿主)→人”的顺序进行环环相扣的迁移。其每一步伴随虫体形态和功能的剧烈转变:
-
虫卵阶段
人体内成熟的肝吸虫可在肝胆管产卵,虫卵随胆汁进入肠道并被排出体外。这些虫卵如果进入水体,可被淡水螺误食,成为新一轮感染的起点。虫卵呈椭圆形,一端有卵盖。 -
螺体阶段分化
在第一中间宿主(如豆螺)的消化道内,虫卵孵化出毛蚴(梨形、具纤毛,可运动),毛蚴穿透肠壁进入组织,转化为胞蚴(囊状、主要进行无性增殖)。胞蚴生成雷蚴(圆柱形,分化出大量后代),雷蚴进一步产生大量尾蚴,而尾蚴则是寄生虫生命周期中极为活跃的传播阶段。 -
鱼体阶段
尾蚴离开螺体后在水中游动,借助其长长的尾巴,寻找第二中间宿主——淡水鱼。当遇到适合的鱼类(如草鱼、青鱼)时,尾蚴穿入鱼体内,失去尾巴,并分泌囊壁形成囊蚴。囊蚴可长期存活于鱼的肌肉组织之中,几个月甚至数年。 -
人体再感染阶段
若人食用了未熟的带囊蚴的淡水鱼(如生鱼片、未煮熟的鱼肉),囊蚴进入消化道,在小肠经胆管进入肝脏,发育为成虫,周期再度循环。
这个复杂的多宿主多阶段发育过程,可以用下表总结相关阶段主要特征和关键变化:
假如有人食用未熟的草鱼片,在人体消化道中的囊蚴即可破囊,幼虫进入肝胆管后生长为成虫,数周后便能产卵,然后虫卵随粪便排出循环再度开始。
防控措施举例:
- 阻断粪便入水,防止虫卵传播。
- 消灭螺类中间宿主,切断虫体发育链。
- 充分加热淡水鱼,杜绝囊蚴感染。
这些都是现实防治中的重点环节,可有效减少感染风险。
华支睾吸虫病在中国南方部分地区仍然流行。生食或半生食淡水鱼是感染的主要途径。预防的关键是改变饮食习惯,确保鱼肉充分加热(中心温度达到63℃以上并持续30秒)后再食用。
通过家蚕的完全变态和寄生虫的多宿主发育我们可以看到,生物体个体在不同发育阶段经历的巨大变化,往往不仅表现在数量的积累,更重在器官的重构、功能的切换乃至生存环境的更替。生命的“同一性”并非固守某个静态特征,而是体现在这一系列动态过程的连续性之中。
细胞代谢的节律性
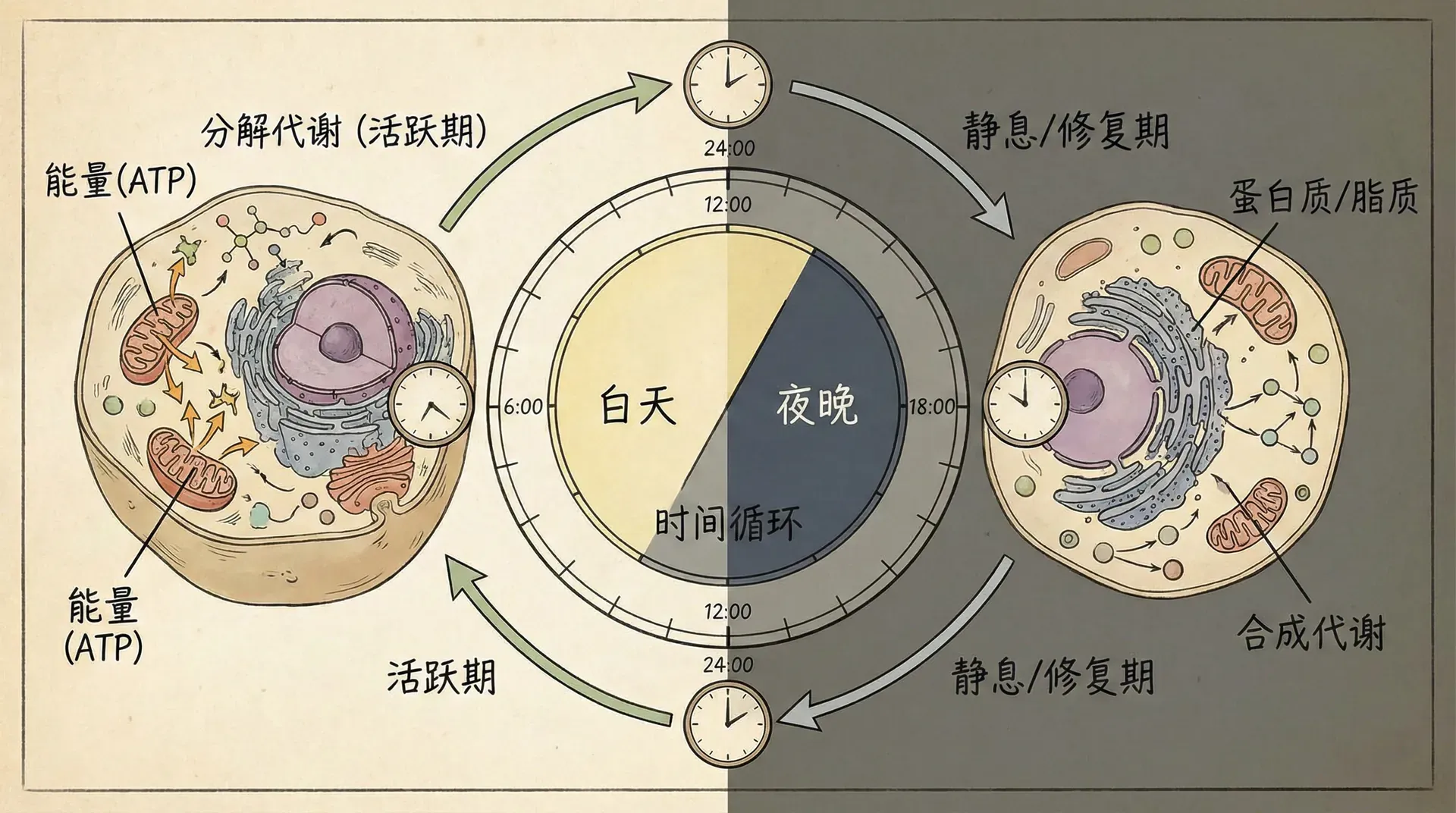
生物体的动态特性不仅表现在生命周期的长尺度上,短时间尺度下,单个细胞的代谢活动同样呈现出规律性的动态变化。研究发现,细胞的生化状态不断随着昼夜节律、代谢周期等因素发生动态切换,并非静止不变。
肝细胞的昼夜代谢节律
肝脏作为人体代谢的核心器官,承担糖、脂、蛋白代谢和解毒等多重任务。分子生物学的进展揭示,肝细胞内许多基因和酶的活动均表现出昼夜节律的波动。例如:
上图展示了肝细胞两类关键代谢酶的昼夜波动。糖原合成酶与糖原分解酶活性的高峰几乎相错,相互切换,确保肝脏能适应机体的能量需求。
肝脏不仅糖代谢如此,脂代谢、胆固醇合成与药物解毒等多条代谢通路也都表现出类似的昼夜节律。例如,细胞色素P450解毒酶家族的活性清晨和傍晚表现出明显差异,这对于药物代谢以及个体对药效和副作用的敏感性具有重要影响。
这种节律性变化的本质是由细胞内的“生物钟基因”调控(如Clock、Bmal1等)。即使在恒定条件下培养,离体肝细胞依然能维持自身的节律。现代“时间药理学”理论据此认为,给药时间的选择可以优化疗效、减少副作用。例如,睡前服用某些降压药比清晨服用疗效更好,这都源于肝脏和全身的节律性变化。
细胞代谢的氧化
不仅仅是昼夜节律,许多细胞还会在几个小时的周期内经历代谢阶段的切换。例如,酵母细胞和部分肝细胞显示出“氧化-还原循环”(Redox cycle),其要点如下:
这种交替,有助于避免氧化应激与合成反应的冲突,提高代谢效率。细胞的节律性和周期性代谢变化,为机体提供了精细的时空调控能力。它不仅有助于维持内稳态,也是适应环境、优化能量利用和应对压力的关键机制。
实际上,不同时间点的同一种细胞,其生理状态与基因表达都可能有极大差异。对医学而言,这揭示了“同一剂量的药物在不同时间给药,效果和风险可能完全不同”的原理。临床中的最佳给药时机选择(如抗肿瘤药、降压药等),往往要结合重要组织如肝脏的代谢节律。
生命的连续性与变化
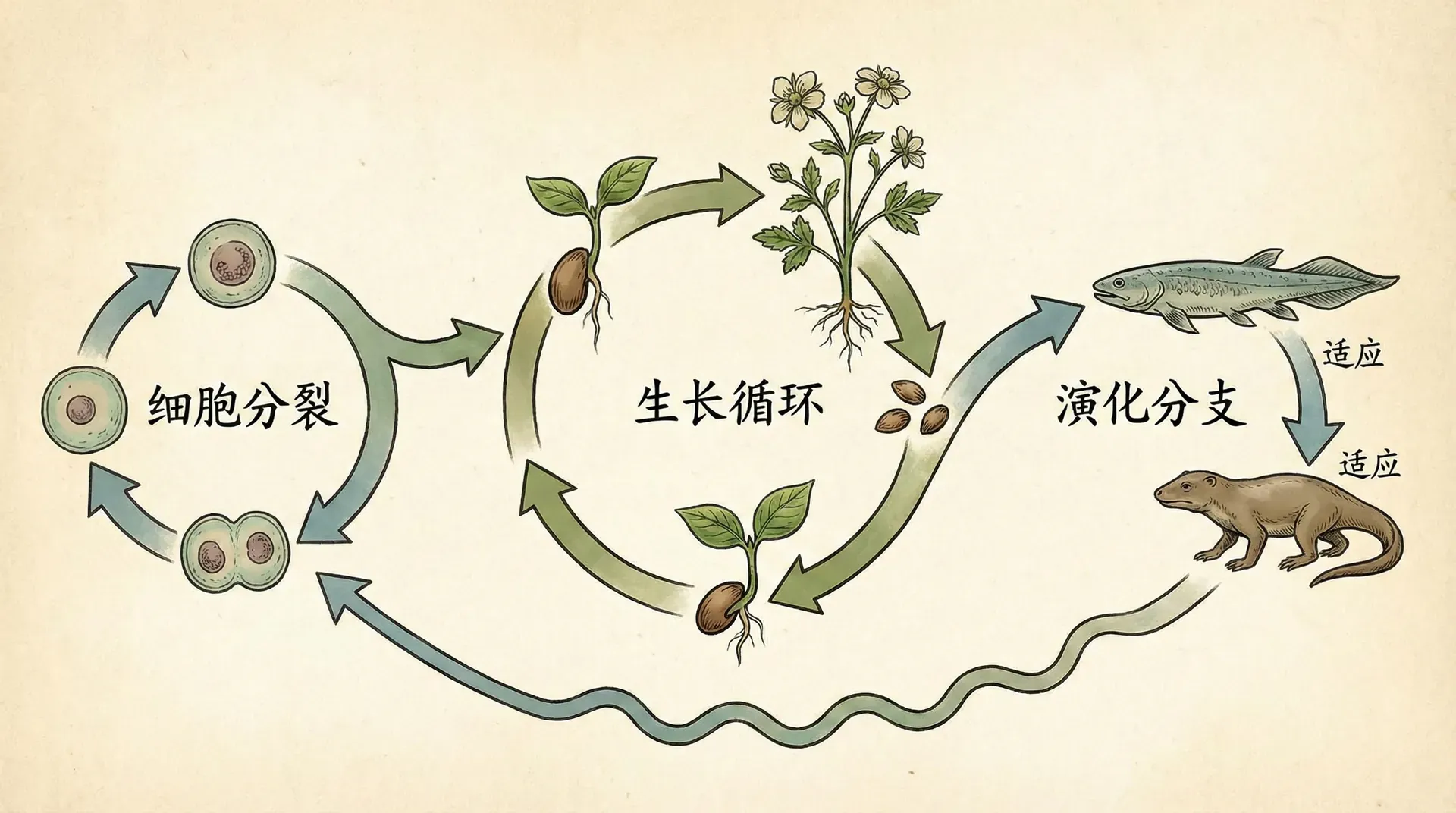
生物体从形态到功能都可以在生命历程中发生巨大变化。对于完全变态的昆虫(如家蚕),甚至各发育阶段外观和生活方式差异巨大:一条蠕动的毛毛虫如何和一只翩翩起舞的蚕蛾被认定为“同一个个体”?
发育
关键在于发育过程的因果连续性:前一阶段为后一阶段提供不可或缺的物质基础与信息支持。以家蚕为例:
虽然每一阶段差异甚大,实际是一个不断延续、递进且依存的链条——你无法跳过任一阶段,也不能割裂地看待整体发育。
这种视角下,个体的同一性并非依赖于始终恒定的形态或分子标志,而是由整个发展过程的连续因果链决定。这类似于河流:无论上中下游多么不同,只要流动未断,就是同一条河。
区分发育与繁殖
理解发育连续性,也帮助我们区分生物体发育(个体更新)与繁殖(个体新生)这两种根本事件。本质区别可归纳为:
- 在家蚕生命周期中,幼虫到成虫是连续发育,物质、结构和信息直接传递;而成虫产卵则产生新的生命体,形成与母体空间分离的个体。
- 以寄生虫为例,华支睾吸虫从毛蚴发育到胞蚴再到雷蚴,是同一生命个体的阶段性转变。而雷蚴无性分裂产生大量尾蚴时,每一个尾蚴本质上已是新的独立体——类似单细胞生物的繁殖。
发育关注连续转变,繁殖则意味着断裂与新生。明辨二者,对于理解物种生活史、疾病传播及种群动态至关重要。
从受精卵到衰老死亡,生物个体始终处于动态的连贯变化之中。正是发育中的因果连续性支撑了“同一生命”的本质。不必执着于找寻某个不变的外在标志——生命之“同一”,源于流变中的不断延续。
理解生物动态过程的意义
理解生命的动态过程,不只是知识体系的革新,更深刻影响着生物学的实践。现代农业与疾病防治等领域中,越来越重视基于生物全周期的信息做出决策。例如,在防控农作物害虫时,仅仅盯住成虫去治理常常收效甚微,因为成虫期往往短暂且个体具备迁飞能力,错过最佳时机就难以遏制虫害扩散。
更科学的做法是分析并预测害虫生命周期的关键节点:如二化螟幼虫孵化初期,尚未钻入稻茎、抵抗力较弱,此刻施药效率最高。这需要将成虫羽化和产卵监测、温度积累与幼虫孵化动态结合,形成完整的过程调控方案。蝗虫治理同理,把握其“蝗蝻期”集中控制,远比等其成虫后大面积扑杀更为高效。
这种思路的本质转变来自动态观的建立。现代生物学越来越强调生命的流动性与过程性。细胞在不断更新组分,个体在发育中持续变化,物种在长时演化中逐步分化、融合——生命不是静止不变的“定格”,而是一系列环环相扣、相互影响的事件流。认识并接受这种动态性,有助于我们跳出只看结构、只比“本质”的静态陷阱,把关注点转向“变化的原因与机制”。于是,细胞、个体与物种的身份,可以在剧烈变化中保持连续性,生命现象的复杂多样性也随之变得更易理解。
把握生命过程的动态本质,不但揭示了生物变化的规律,也为实际应用打开了新思路。从精准的病虫害管理,到疾病的发生发展机制、再到生物技术的创新突破,动态视角是理解和利用生命的关键。
归根结底,生命世界如一场绵延不断的交响乐,由无数过程交融、推进。只有理解并顺应这种动态特性,我们才能真正理解生命、有效应用生物学知识,推动科学与社会的进步。