表观遗传学
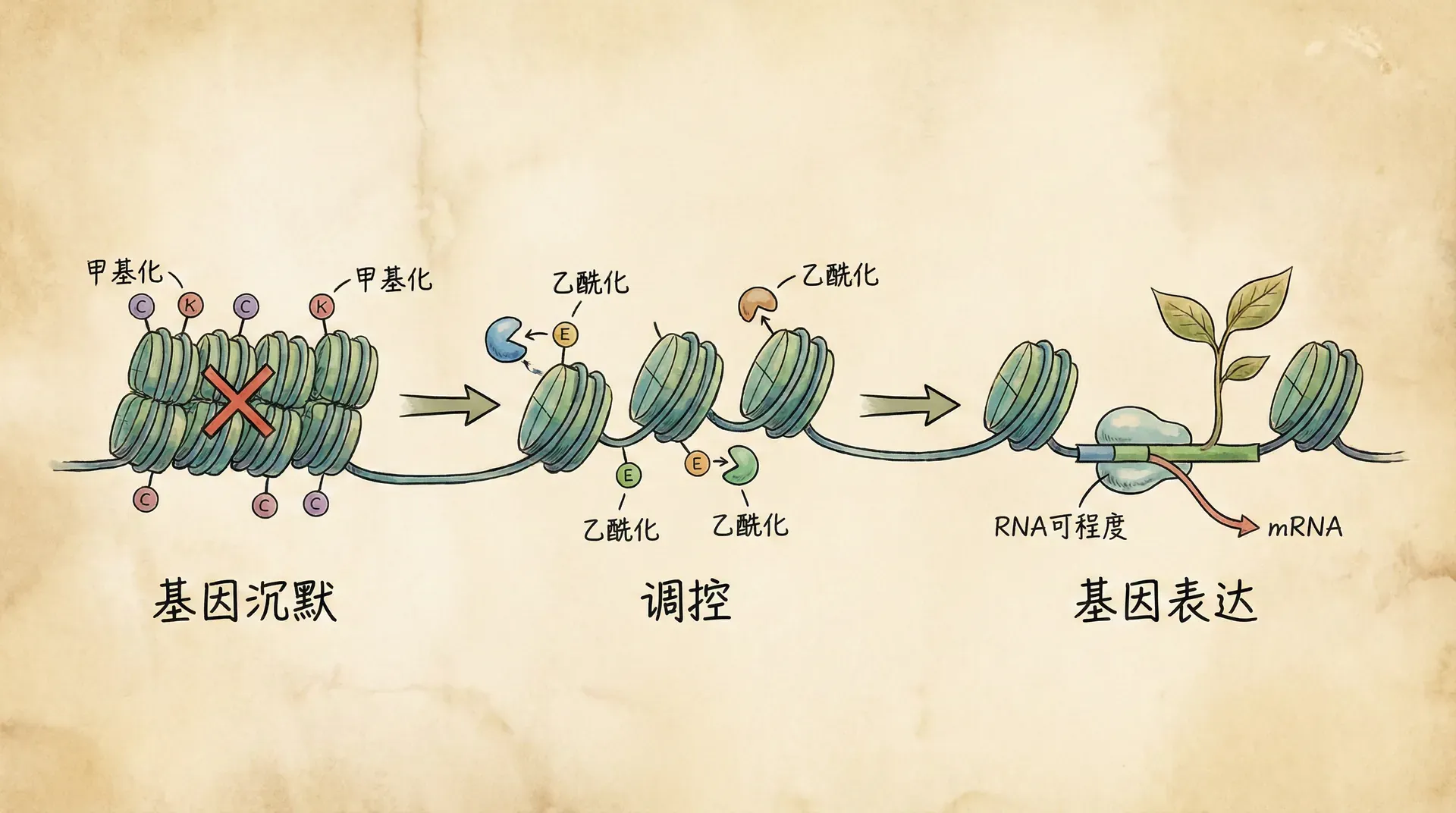
大熊猫的祖先原本是食肉动物,但如今它们却以竹子为主食。这种巨大的食性转变并非经历了数百万年的缓慢基因突变,而是在相对较短的时间内完成的。再看看生活在青藏高原的藏族同胞,他们能在海拔四千多米的高原上自如生活,而平原地区的人初到高原往往会出现严重的高原反应。这种适应能力的差异,背后隐藏着怎样的生物学机制?
传统的遗传学告诉我们,生物的性状由基因决定,进化通过基因突变和自然选择实现。但这个解释面临一个难题:基因突变是随机且缓慢的,而生物却常常能在短时间内对环境变化做出快速响应。这就引出了一个关键问题——生物如何在环境压力下快速适应?仅仅依靠基因突变真的够吗?
答案就藏在表观遗传学(Epigenetics)这个领域中。表观遗传学研究的是,在不改变DNA序列的前提下,生物如何通过调控基因的表达方式来改变自身的性状。这就好比一本菜谱,基因是写在书上的食谱,而表观遗传机制则决定了今天翻到哪一页、做哪道菜。同样的基因“菜谱”,在不同的环境条件下,可以“烹饪”出截然不同的生物性状。
在20世纪40年代,英国发育生物学家沃丁顿(Conrad Waddington)就提出了一个革命性的观点:发育过程不是基因程序的简单执行,而是一个动态的、具有调控能力的过程。
生物体就像一个智能系统,能够感知环境变化,调整自己的发育路径,甚至将这些调整传递给后代。这个观点在当时颇具争议,但如今越来越多的研究证实了他的远见。
发育的稳定性与可塑性
在理解表观遗传学之前,我们需要认识发育过程的两个看似矛盾的特征:稳健性(robustness)和可塑性(plasticity)。稳健性是指生物抵抗环境干扰、保持自身特征的能力;而可塑性则是指生物根据环境变化调整自身性状的能力。这两个特征听起来相互矛盾,但实际上它们是生物生存的两个方面。
中国是世界上最大的水稻种植国,从黑龙江到海南岛,从沿海到高原,水稻品种展现出了惊人的多样性。北方的粳稻能在较低温度下正常生长,而南方的籼稻则适应高温多湿的环境。这种差异背后,就是稳健性和可塑性的平衡。
一个水稻品种如果完全没有稳健性,那么温度稍有波动就会导致发育异常,无法形成稳定的产量。但如果完全没有可塑性,那么这个品种就只能在非常狭窄的环境条件下生长,一旦遇到干旱、高温或低温就会绝收。优秀的水稻品种需要在这两者之间找到平衡点:既能在常规条件下稳定生长(稳健性),又能在极端天气来临时调整生长策略以求生存(可塑性)。
家蚕是中国古代四大发明之外的又一伟大成就,经过数千年的人工驯化,野生桑蚕变成了如今完全依赖人类照料的家蚕。这个驯化过程中,人类实际上是在调整家蚕的稳健性和可塑性平衡:增强了对人工养殖环境的稳健适应(比如对密集饲养的耐受),同时保留了对桑叶质量变化的可塑性(能够在不同品质的桑叶条件下调整生长速度)。
从上图可以看出,高稳健品种在低到中等压力下表现出较小的表型变化,但一旦压力超过阈值就会快速失控;而高可塑品种从一开始就展现出较强的响应能力,能够渐进式地调整自己的表型,但在极端压力下的变异程度略小于高稳健品种崩溃后的状态。这说明,不同的生存策略适应不同的环境条件。
那么,生物是如何同时实现稳健性和可塑性的呢?答案在于发育过程的动态调控机制。在细胞层面,基因和细胞质之间存在着复杂的反馈循环。基因不是单向地“指挥”细胞发育,而是与细胞质环境进行持续的“对话”。当环境条件发生变化时,细胞质会捕捉到这些信号,并通过反馈机制影响基因的表达模式。这种双向互动保证了发育过程既有章可循(稳健性),又能随机应变(可塑性)。
同态流
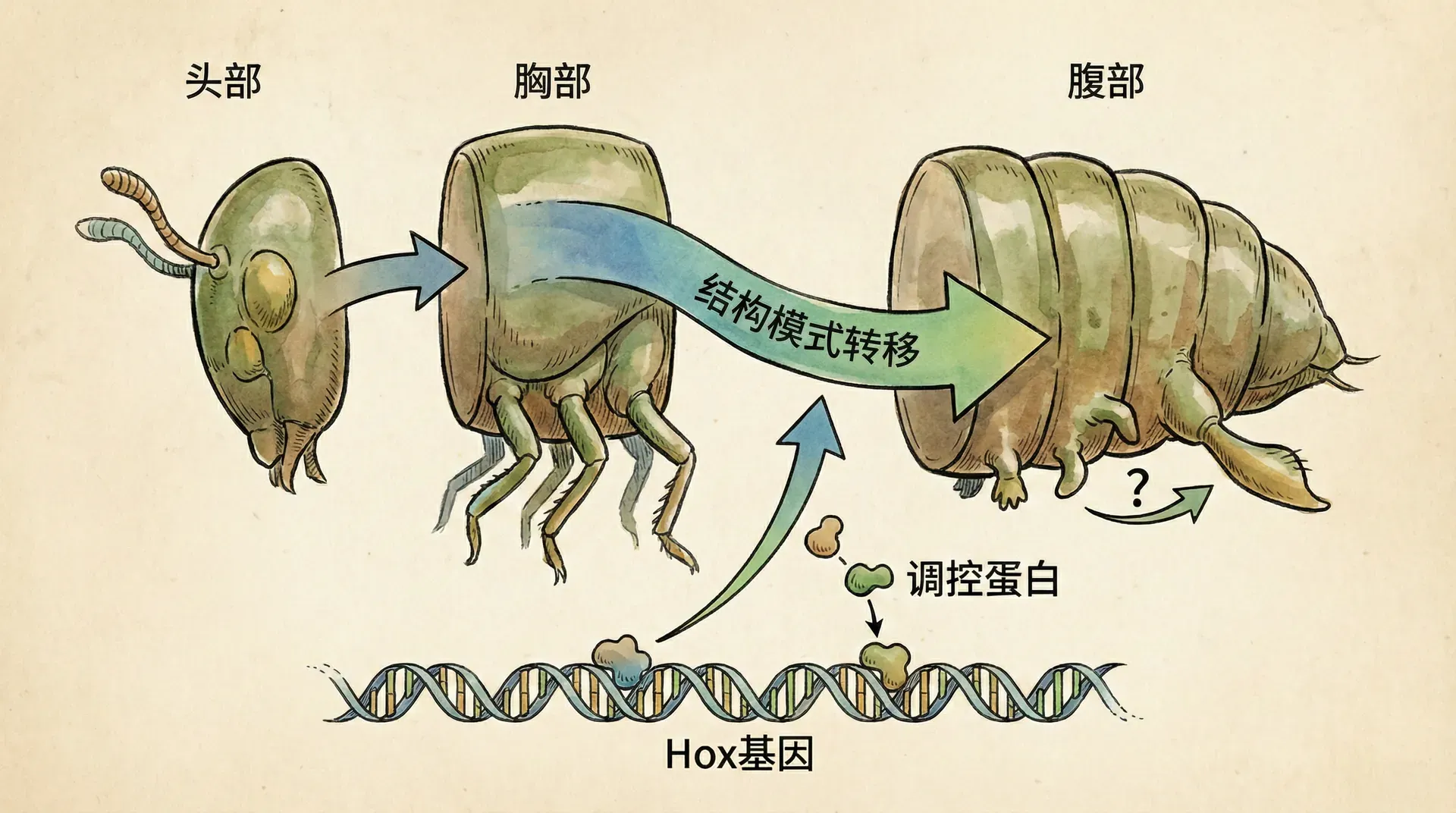
为了更准确地描述发育过程中稳健性和可塑性的统一,沃丁顿提出了一个重要概念——同态流(homeorhesis)。这个概念常常与我们熟悉的稳态(homeostasis)混淆,但两者有着本质的区别。
稳态描述的是系统维持某个固定状态的能力。比如人体的体温调节就是典型的稳态机制:无论外界温度如何变化,人体总是试图将核心体温维持在37℃左右。这是一个“点对点”的调控,目标是回到某个特定的数值。而同态流描述的则是系统维持一个动态变化轨迹的能力。用一个形象的比喻:稳态就像把一个球固定在一个坑里,无论怎么推它,它都会滚回原位;而同态流就像让一个球沿着特定的轨道滚下山坡,无论中途遇到什么小的干扰,它总能回到预定的轨道上。
发育过程本质上是一个同态流过程。一个受精卵发育成完整生物体,需要经历一系列精确编排的步骤。在这个过程中,发育路径会不断分叉——这就是沃丁顿著名的“表观遗传景观”模型所描述的情景。想象一座山顶上的一个球,山坡上有许多分叉的沟壑,球滚下来时会在每个分叉点选择一条路径,最终到达山脚的某个位置。这个球代表一个正在发育的细胞,沟壑代表可能的发育路径,山脚的位置代表不同的细胞类型(如肌肉细胞、神经细胞等)。
中国是鲤鱼养殖的起源地,几千年的养殖历史中积累了丰富的经验。研究发现,同一批鲤鱼卵,如果在不同的养殖条件下孵化和生长,会发育出不同的体型特征。在高密度、高投喂的池塘中生长的鲤鱼,会发育出相对短粗的体型和发达的消化系统;而在低密度、自然觅食的湖泊中生长的鲤鱼,则发育出流线型的体型和强健的游泳肌肉。
从图中可以看出,虽然不同环境条件下的鲤鱼发育速度和最终的分化程度略有差异,但它们都遵循着S型曲线的基本模式:早期发育较慢,中期快速分化,后期趋于稳定。这就是同态流的体现——无论环境如何变化,发育的基本轨迹保持稳定,只是在轨迹的细节上做出调整。
这种发育路径的渐进限制被称为“渠化”(canalization)。在发育早期,细胞具有广泛的分化潜能,可以成为多种类型的细胞;但随着发育的进行,这种潜能逐渐收窄,最终被“锁定”在特定的细胞类型上。这就像水流从宽阔的河口进入狭窄的河道,最初可以向多个方向流动,但最终会被地形约束在特定的路径上。渠化保证了发育的稳健性,避免了随机因素导致的混乱;同时,多条并存的“河道”又提供了可塑性,使生物能够根据环境选择最合适的发育路径。
同态流机制揭示了一个重要原理:生物的稳定性不是静止的,而是动态的。发育过程不是机械地执行遗传程序,而是在遗传信息的指导下,根据环境反馈不断调整的动态过程。这种动态稳定性使生物既能保持物种的基本特征,又能适应环境的多样性。
获得性状的遗传

在观察不同生物适应环境的过程中,我们常常会思考:环境诱导的性状改变是否有可能遗传给后代?长期以来,传统的达尔文进化论和孟德尔遗传学都坚持“获得性状不能遗传”的观点,认为只有基因突变和重组等内源性变化才可遗传。然而,沃丁顿(Waddington)在20世纪中期的实验却揭示了一个更复杂的现象:在特定条件下,最初由环境诱导产生的性状,可以经过多代筛选,变成无需环境诱导即可稳定表达的遗传性状。 这一过程,被称为“遗传同化”(genetic assimilation)。
遗传同化实现的关键在于“隐性变异”。平时,生物体内存在着丰富的遗传变异,但它们很少表现出来,原因是发育系统有强大的稳健机制来“隐藏”这些变异。当环境压力增大到一定程度后,这些被掩盖的变异就有机会浮现并表现为新的表型。如果这些新表型有助于个体适应新环境,那么自然选择会促使这些变异的频率逐代升高。最终,即使最初的诱发性环境消失,这些特征也能自发地、稳定地遗传下去。
以下对比“传统遗传”与“遗传同化”的逻辑:
案例举隅
-
果蝇的交叉脉实验
沃丁顿早期通过高温处理果蝇幼虫,诱导部分个体翅膀出现“交叉脉”异常。数代选择后,这一异常在未加高温的后代里也能自发表达,表明原本需要环境诱导的性状已经转为基因控制。 -
金鱼的演化历史
金鱼原为鲫鱼的野生种,早期的奇异体色多是池塘等特殊环境诱发的“突发变异”。人工筛选后,类似龙睛、绒球等形态性状,成为无需特殊环境即可遗传稳定表达的家养品系。 -
西藏高原人群的高原适应
西藏人群的高原适应性是另一个经典例证。初期进驻青藏高原的人祖先,通过暂时增加红细胞等应激性反应度过低氧压力。随着EPAS1等基因变异的积累、筛选,后代表现出无需额外增加红细胞即能高效携氧的遗传适应。下表总结该案例关键点:
上图展示了遗传同化的动态过程。在持续环境压力下,有利性状在群体中的比例逐渐提升,最终几乎全部个体都具备该性状;而如果中途移除环境压力,则提升趋势减缓或停止。由此可见,遗传同化是自然选择和发育可塑性共同作用的产物,而非“用进废退”式的简单获得性状遗传。
遗传同化发生的关键基础是发育系统的缓冲能力。平时,同态流等机制维持了表型稳定,但当累积压力超出缓冲上线后,隐藏变异被释放、获得性状频率迅速提升。原本需要外部刺激的表型逐渐转变为无需环境即能表达的遗传性状,进化速度大幅提升。
遗传同化不仅深化了我们对达尔文自然选择理论的理解,还揭示了环境能够“激活”潜在的遗传多样性,为自然选择提供更多素材——进化不仅是被动的筛选,还包括潜能的主动释放与塑造。
隐性变异
讨论遗传同化,就一定绕不开“隐性变异”的来源。它是长期隐匿于基因组的“库存”,还是压力下实时“制造”出来的?现代生物学对此提出两大机制模型,每种都有经典案例和代表性分子基础。
模型一:缓冲释放模型
该模型认为,生物体长期积累了大量潜在的遗传变异,但在正常情况下,它们被分子伴侣等机制有效“隐藏”。典型代表分子——热休克蛋白Hsp90,可帮助各种蛋白折叠修复,掩盖不利突变的表型效应。只有在强烈环境胁迫(如高温、药物等)下,Hsp90功能受损,变异效应突然释放,表现为形态或功能异常。
这一过程如同“水库大坝”模式:
- 平常:大坝拦蓄隐性变异,个体表型稳定。
- 胁迫来临:大坝溢流,杂多变异一拥而出,表现为突变性状激增。
模型二:动态产生模型
与上者不同,动态产生模型强调新变异是在环境压力下“制造”出来的。其核心分子为“转座子”——可以在基因组中跳跃移动的DNA片段。压力状态下,原本受抑制的转座子被激活,通过插入或重排基因组,快速制造大量新突变。部分新变异虽有害,但也可能供进化利用,成为适应新环境的原材料。
以下对比两种模型的主要特性:
- 果蝇Hsp90实验:科学家用药物抑制Hsp90后,果蝇机体出现多种异常表型,杂交选择数代后,这些表型变成无需化学抑制即可遗传。
- 植物抗逆适应:在植物如阿拉伯芥的干旱胁迫实验中,发现转座子激活后,部分后代引入了新的基因结构变异,部分变异可被稳定遗传,增强了抗逆能力。
图中,缓冲释放方式的遗传变异量初期迅速上升后趋于平台,动态产生方式则可持续累积更多新变异,进化和适应能力更强。在现实自然界中,如玉米、阿拉伯芥等多种植物长期面临环境胁迫,常常呈现变异长期持续增加的趋势,这支持了“动态产生”机制在适应性进化中的核心作用。
需要强调的是,两种机制并非互斥,实际上在许多生物中它们都可能共存。例如,动物急速适应环境时可能先释放隐性变异(缓冲释放),后续则通过转座子等机制持续产生新的遗传创新(动态产生),共同保障适应速度和进化潜力。这种“应急-创新双保险”是生命系统抗风险、促多样化的关键策略。
转座子曾被误认为“垃圾DNA”,但现在已知是基因组重塑和适应性创新的重要引擎。在生命系统的复杂调控网络中,许多表面无用的元素往往潜藏着巨大的演化价值,我们应以更开放的眼光重新审视它们的意义。
总结
自沃丁顿提出表观遗传学以来,这一领域已取得诸多进展,并在农业、医学、生态保护等领域展现出切实价值。
在农业中,表观遗传学突破了传统育种的瓶颈。通过特定环境处理可激活作物隐藏变异,加快培育适应性新品种的速度,有助于应对气候变化。在医学领域,表观遗传机制揭示了疾病的环境根源。例如,孕期营养状况影响后代代谢健康,强调了早期干预和疾病预防的重要性。
在生态保护方面,表观遗传学提供了预测和提升濒危物种适应力的新思路,不仅仅考量基因多样性,更关注发育过程的可塑性,为小种群保护带来希望。实际上,许多珍稀动物(如圈养大熊猫)在面临环境变化时,关键在于调控其发育敏感期,模拟自然刺激,以促进适应性特征的形成。
表观遗传学告诉我们,生命不仅仅是基因的“程序”,更展现出因环境而变、主动塑造自身的能力。理解和利用这种可塑性,将推进农业生产、健康管理和生物保护的创新发展。
总之,表观遗传学不是否定基因作用,而是完善我们对生命和进化的整体认知。基因、发育与环境相互作用,决定了生命的多样与韧性。只有整合这三者,我们才能真正把握生物学的核心奥秘,更好地服务于现实挑战。