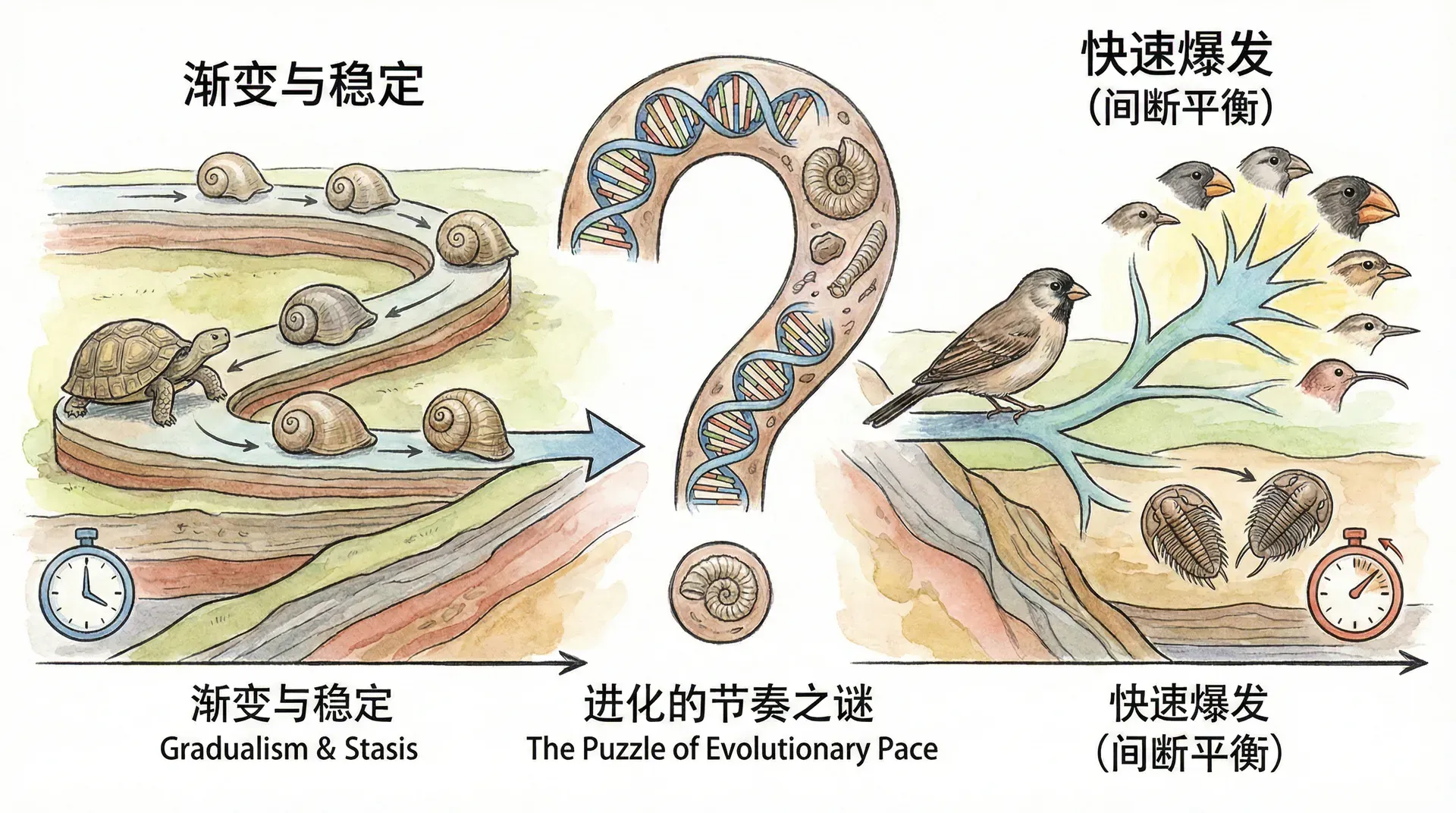
生物进化的节奏之谜
在古代丝绸之路上,商队从长安到罗马,跨越7000公里的遥远旅程。这条道路不仅是东西方经济与文化的纽带,也是充满挑战的漫长征途。如果按照出发和到达时间简单计算其“平均速度”,会得出商队每天只前进约19公里的结果,甚至比普通步行还慢,显然与我们对驼铃悠悠、日夜兼程的想象不符。
实际上,商队的行进模式远比平均数复杂得多。他们会在荒漠与戈壁中快速穿行,争分夺秒赶路,只为在水源有限的困难地带及时抵达绿洲。而每到一个绿洲,商队则会长时间停留,补充补给,修整行装,甚至因各种突发原因(如天气、边境动荡、贸易商议等)被迫滞留数周乃至数月。每一次完成休整再出发,往往又是高效而迅速的“冲刺”。因此,丝绸之路的实际行进轨迹,是由频繁的“停顿”与间或的“快进”交错构成,呈现出“阶段性停滞-短暂冲刺”的节奏。只看整体平均,反而遮蔽了这其中的动态细节与独特节律。
同样道理,假如一个考察者只关注商队始终点之间的平均速度,必然会忽略中间复杂的阶段与细节,这与我们根据化石记录仅凭“间断”来判断生物历史有着异曲同工之处。
这就引出了生物进化中的一个核心争议:生物进化到底是像钟表一样匀速进行,还是像丝绸之路商队那样,经历长期的静止和偶发的快速变化?生物的历史是一条平滑上升的曲线,还是一连串“停滞-突变-再停滞”的阶梯?
间断平衡论
1972年,美国古生物学家尼尔斯·埃尔德里奇(Niles Eldredge)和斯蒂芬·杰伊·古尔德(Stephen Jay Gould)首次系统提出了“间断平衡论”(Punctuated Equilibria)。他们仔细研究了丰富的化石资料,发现绝大多数物种在很长的地质时间内基本保持形态稳定,只有在较短的时间窗口里才会发生较大的进化变化。这一理论的核心观点可以用三个关键词概括:
这个理论形象地揭示了进化史中的两个世界:一个缓慢、稳定、几乎原地踏步;另一个则在某些时期急速激变,走完了常人想象中几万年甚至几百万年才能完成的演化之路。许多地质记录都能看到这种“阶梯式”的变化轨迹。下表和图示进一步阐释了这两种进化观的差别:
与传统观点的区别
间断平衡论并不否认达尔文的渐进进化思想(gradualism),也不是说进化“存在飞跃”,而是强调进化速度在时间上分布不均,而非线性渐变。传统观点认为生物进化是一个持续、缓慢累积的过程,物种形态沿着“平滑斜坡”不断变化和完善;而间断平衡论基于大量化石与地质证据,提出物种在漫长时期内基本保持稳定,仅在特定的环境剧变、迁徙或小种群隔离等关键节点才发生较快的进化“跃迁”,整体表现为“静止-突变-再静止”式的阶梯变化。因此,进化节奏真正的特点,是大量静止阶段夹杂短暂的快速变化期,这种不均匀与生态、遗传、地理事件密切相关。
需要注意的是,这里的“间断”并非指进化中有突然的、超自然的飞跃或颠覆性变化。即使在快速变化期,物种演化依旧依赖代际间自然选择与遗传变异的逐步积累。只不过在地质时间尺度下,这些累积被压缩进更短区间,在化石记录中给人“突然出现新类型”的印象。实质上,这种“突变”依然符合生物学规律和自然法则。
中国生物多样性的证据
中国地域辽阔、地形多样,拥有丰富的自然生态系统,这使得中国不仅成为世界生物多样性最为丰富的国家之一,也为研究进化过程特别是间断平衡论提供了宝贵的天然样本。中国的生物多样性在植物、动物、真菌等多个门类上均表现出极为突出的特征,许多“活化石”、地理隔离现象与快速物种分化的经典实例,都成为验证和拓展进化理论的真实案例。
活化石现象
中国拥有全球数量众多、种类齐全的“活化石”资源,这些古老生物代表着生命演化史上的原始形态,对理解进化的“静态期”具有重要意义。其典型物种包括:
从表中可以看出,这些物种自古至今在化石记录中形态变化极小,展现出异常悠久的“静态期”,成为间断平衡论下的典型“活化石”代表。
这些活化石不仅数量丰富,而且往往具有极强的环境适应性。例如:
- 银杏的启示:银杏树被称为“植物界的活化石”,叶片形态从2.5亿年前到今天基本完全一致。这种惊人稳定性,说明在长期相对稳定的生态环境下,自然选择更倾向于保留现有适应性特征,从而延长了“静态期”。自二叠纪以来,银杏经历了多次冰期、气候变迁,但因其适应了多种环境、具备较强耐寒抗旱能力,很少被外界激烈变动淘汰。
- 扬子鳄与中华鲟:作为生活在长江流域的古老动物,它们在形态和生态行为上都和史前祖先惊人接近。中华鲟现存种群与2亿年前的化石相比,头骨结构几无变化,是“进化缓慢”的生动样本。相反地,这类物种的数量在现今环境压力加剧时急剧减少,显示其“静态”难以应对剧烈社会变化。
这些活化石的存在表明,在合适的环境条件下,生物可以在极长的时间内保持形态稳定,这正是间断平衡论中“静态”概念最好的现实注脚。它们映证了进化并非始终是连续渐变,而是阶段性地高原期和突变期相间的过程。
地理隔离与物种快速分化
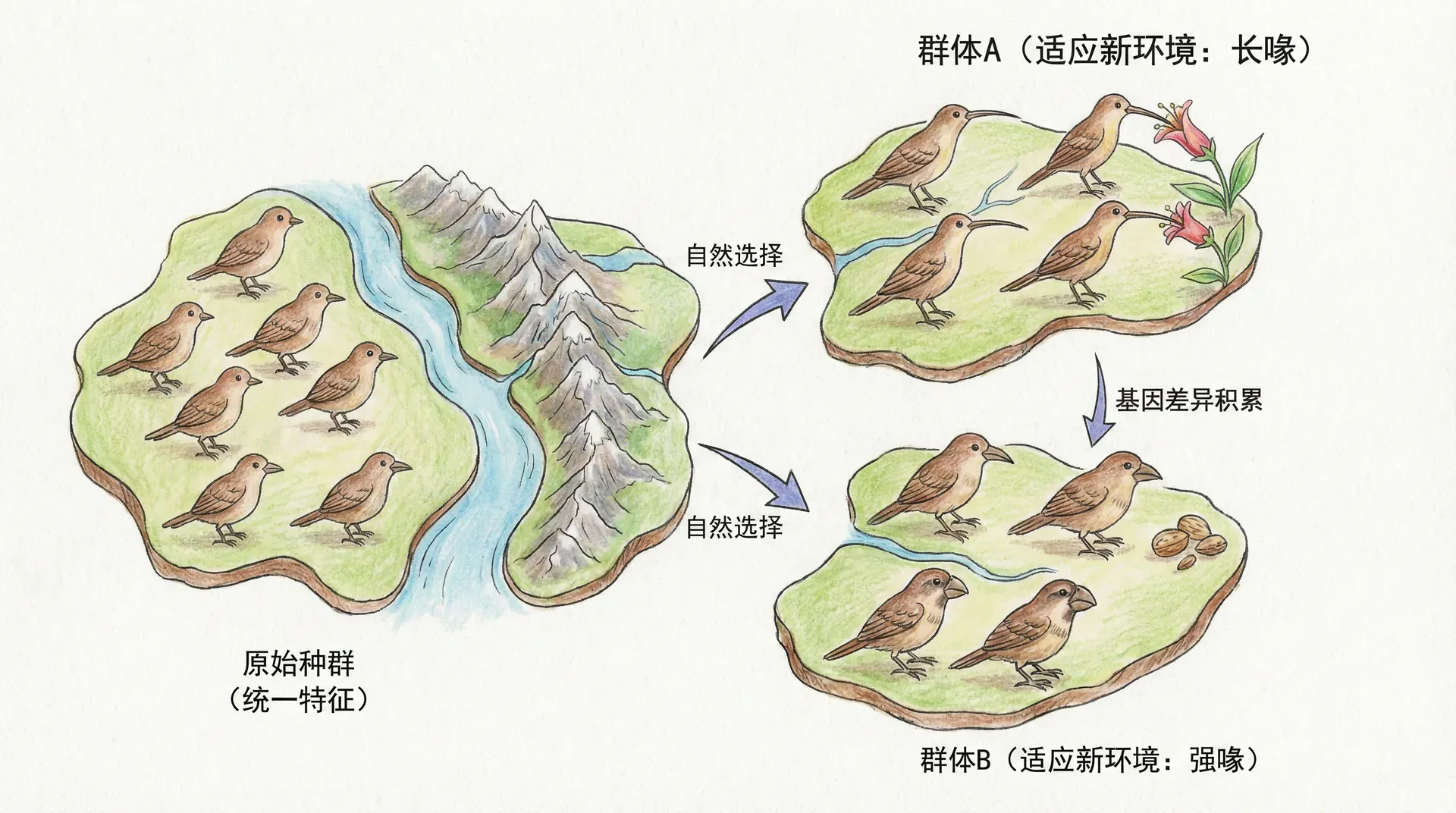
中国复杂的地形地貌(高山、河谷、盆地、荒漠)为生物的地理分化提供了天然平台。不仅秦岭-淮河线分隔了南北气候生态,更有青藏高原、横断山脉、台地河谷等形成独特生态屏障。不少物种正是在这些地理隔离条件下实现了快速分化,成为间断平衡论下“间断—平衡”机制的优质案例。
大熊猫的种群分化
大熊猫作为中国的“国宝”,其种群分化历史堪称间断平衡论的标准范式。约30万年前,秦岭山脉的快速隆起将大熊猫祖先种群分割,形成了秦岭亚种和四川亚种。两地海拔、气候、植被差异显著,且隔离之后环境波动剧烈,外部选择压力加大,大熊猫便在短时期内迅速沿不同方向分化。
大熊猫的分化过程揭示了间断平衡论的三个核心机制:
- 地理隔离的建立:秦岭的隆起在短时间内形成了强隔离屏障,阻断基因交流。
- 快速分化期:隔离后10~15万年内,受到气候和植被差异影响,两地种群迅速呈现遗传与形态差异。例如,秦岭大熊猫体型更小、头骨更圆、毛发密度更高,适应山地多云湿冷环境;四川大熊猫则体型稍大,更适于亚热带密林生存。
- 新的平衡状态:分化完成后,两亚种进入新的相对“静态期”,各自稳定适应环境。
现代基因组测序证实,秦岭大熊猫与四川大熊猫的基因结构、头骨形态、免疫抵抗力等多方面存在显著差异。这种分化在十几万年间完成,正好符合间断平衡论“快速分化—新平衡”的理论预测。
同时,中国境内还有许多类似案例:比如金丝猴(滇金丝猴、川金丝猴)、朱鹮种群等在横断山脉与秦岭区域的分化历史,都有力说明了地理隔离在推动快速进化中的作用。
青藏高原

青藏高原作为世界上海拔最高、地质变化最为剧烈的区域,被称为“第三极”,封存着许多生物进化奇迹。约300万年前开始的青藏高原快速隆起,不仅塑造了极端的气候和生态条件,也大幅促进了我国高原物种的分化和适应,成为间断平衡论实地观测的绝佳窗口。
藏羚羊的高原适应革命:
藏羚羊祖先原本生活在较低海拔,但随着高原升高,这一部分族群被“抬升”到恶劣低氧、低温、高紫外线的极端环境,短时间内演化出多项适应性“创新”:
在这一过程中,基因组检测显示,藏羚羊的耐高原、耐缺氧遗传位点(如HBB、EPAS1等)都发生了快速的扩散,体现出“突变—适应—定向选择”在短时间内完成进化的节奏。这些适应性特征的出现与青藏高原的地质剧变高度同步,成为“环境剧变—进化加速—新平衡”理论的重要实证。
不仅限于藏羚羊,藏马鸡、高原鼠兔、雪豹等青藏高原特有物种也都表现出类似“高原适应性跳跃”现象。大量研究显示,高原的快速隆起及破碎地形加剧了物种隔离和生态位分化,大批动植物在数万年至十万年间出现了爆发性物种形成,这一事实与间断平衡论高度契合。
理论争议与科学证据
不同进化速度理论的对比
进化生物学关于进化节奏主要有以下理论流派,不同理论在中国实际观测中都有案例呼应:
这些理论往往并非绝对对立,而是在不同物种、不同生态压力、不同时间尺度和遗传层级上各有适用。中国繁复的生态与进化历史,为多种理论的比较研究提供了理想平台。
人工选择实验的启示
中国农业育种历史悠久,人工选择的实验对理解进化速度和模式至关重要。相比许多西方经典案例,中国丰富的农作物和家养动植物育种史是检验间断平衡论的宝库。
杂交水稻的中国奇迹:
袁隆平院士领导的杂交水稻研究,是人工选择导致生物特性的快速变异与“新平衡”的典型。
- 快速响应:强选择压力下,水稻产量、抗倒伏、抗病等性状在短短几代内显著提升,表现为阶段性跳跃式变化而非渐变。
- 阈值与突破:经过反复杂交优选,某一特定遗传组合达到“阈值”,性状发生明显分化,实现“突变”式提升。
- 新稳定期:突破后新品种表现出较强遗传稳定性,可以连续代次遗传,体现为“新平衡期”。
茶树的地方性分化:
中国多样的地貌和气候孕育出龙井、铁观音、普洱等极具地方特色的茶树品种。这种品种分化往往在若干十年或百年内发生,且性状分化阶段性强,具有鲜明的“间断性”特征。例如:
- 龙井茶只在杭州一带形成特有茶多酚比值、香味物质;
- 普洱则发展出耐高温、耐湿热的遗传特征;
- 铁观音发源于安溪,遗传多样性极高,展现出地方性多样化非常典型的“分步进化—稳定—再进化”分化模式。
这些快速变异和品种的固化,说明在人工和环境压力耦合作用下,生物性状的改变可以远比传统“渐变论”预期更快、更分阶段。
现代分子证据的佐证
分子生物学的快速进步带来了全新视角,推动我们从基因、蛋白质、基因组调控等底层机制验证进化速度理论。
因此,现代中国生物多样性的诸多“奇观”,无论在古生态化石、地理隔离、人工选择还是分子生物学层面,都为间断平衡论的理论体系提供了丰富而生动的本土证据。这些实例不仅加深了我们对进化节奏复杂性的认识,也让中国成为全球研究多样性和进化模式的重要前沿。
误解与澄清
常见的误解
间断平衡论自提出以来,经常被误解、简化或者过度解读,甚至在科学传播和大众科普中流传着许多似是而非的观点。以下是一些在学术界和公众领域中广泛存在、需要特别加以澄清的主要误区:
其他常见误区:
- 有些人以为间断平衡论只适用于“高等动物”,但实际上它也可用于解释某些植物、微生物甚至病毒的快速适应过程。
- 还有人将“快速变化”等同于“基因突变剧增”,但间断平衡期间的变异主要来自积累的隐性遗传变异以及环境敏感性,并非突变率极大升高。
科学争论的价值
间断平衡论的提出,一度在进化生物学界引发了长期、激烈甚至是带有个人色彩的学术争论。从70年代首次提出至今,关于物种形成速率、化石记录可信度、遗传机制解释、统计方法等方面,学界产生了丰富而细致的讨论。这种争论的过程深刻体现了科学发展的动力机制:
建设性的争论推动了理论创新和研究工具的进步:
- 促进了对化石记录时空分辨率的再思考,以及对“化石间断性”的多角度解释;
- 推动了进化速率的新数学模型发展,包括分支点过程、变速模型、计算流行遗传学的应用;
- 激发了跨学科合作,例如结合分子生物学与古生物学、统计物理方法等,从更细致的尺度检验和拓展理论边界;
- 鼓励了长时间尺度、高时间分辨率的实验设计,以及野外-实验室联合、纵向队列等多样化研究范式。
正因为有争论、质疑、补充,科学才不断迈向更真实和全面的理解。
总结
回到开头的“丝绸之路”类比:商队在长途跋涉中,大多时间缓慢前进、积蓄力量,在关键节点(如山口、荒漠、河谷)则抓住机遇,实现快速跃升。生命的演化节奏也同样如此——大多数时期内,物种在稳定环境下缓慢演化、保存特性;而每当地质、气候或人类活动等外部事件出现时,又会成为创新与跃迁的催化剂,如同一阵风暴或一条新商路,推动生命在“静与动”“守与变”之间持续前行。
实际上,间断平衡论自身的发展史也是科学进化的缩影:自1972年提出以来,经历了激烈争论与大量证据的积累,不断与分子生物学、古生物学、生态学等领域成果交融,拓展和完善了我们对生命演化速度与节奏的理解。
如今,间断平衡论已深深融入现代进化科学体系。它提醒我们,生命历史是停滞与爆发、平稳与跃进循环交替的过程。理解这种节奏,不仅有助于洞察多样性的根源,也为生物多样性保护与人类应对未来环境挑战提供了新的视野和方法。
如同丝绸之路串联起世界各地,生命亦在“间断与平衡”的节奏中创造出今日的繁盛景观。这种进化的智慧——在静待与突破之间积蓄、创新——正是自然界赋予我们的宝贵财富,也是我们不断理解与改造世界的灵感源泉。