生物分类学的奥秘
生物分类学乍一看仿佛枯燥又无趣,很多人会将它和记忆复杂名称、死记硬背枯燥知识联系在一起。但事实上,分类学远不止如此——它是揭示生命世界奥秘的“钥匙”,帮助我们理解地球亿万年生命演化的宏大图景。想象一下,正如考古学家能够凭借几片陶器残片还原出古代文明的面貌,生物分类学家则通过分析现生与化石生物的各种特征、行为、分子信息,重建出生命演化的历史脉络。从细胞的演化到脊椎动物的崭露头角,再到人类自身出现,每一步都刻画在分类系统中。
而在中国辽阔多元的地理环境下,从皑皑雪山的珍稀雪豹、青藏高原的藏羚羊、到华北平原上渺小却顽强的草履虫,每一种生物都是这幅进化画卷上的一笔。它们彼此间千丝万缕的联系,不仅映射着生物多样性本身,也蕴含着生命起源与演化的深层秘密,等待我们一步步去揭开、去理解。
为什么需要给生物分类
从整理图书馆说起
如果你走进一个藏书丰富的大型图书馆,里面有数百万本书籍,但却没有任何分类系统或索引,你会是什么感受?想象一下此时你需要查找一本关于大熊猫研究的书,却发现生物学教材、民间故事小说、菜谱、诗集、物理学论文全都被随意混杂在一起,没有章法。面对如此混乱的环境,即使拥有丰富馆藏,读者也只能望洋兴叹。使用和管理都变得极其低效。
为了解决这个问题,图书馆管理员会设计并实施一套分类体系。按照学科门类分区、根据出版年代分组、甚至以作者姓氏音序排列,都可以让信息变得井然有序。只要分类标准清晰、索引准确,书籍的管理和查找效率便大大提升。不同图书馆可能采用不同方案:有的更注重学科,有的侧重地域与历史时期。例如中国国家图书馆采用《中国图书馆分类法》,美国国会图书馆则使用LC分类法。这些系统互不相同,却各自运转良好,满足了不同用户的需求。
值得注意的是,这里存在一个根本特征——对于图书分类,没有所谓唯一的“标准答案”。分类标准是“人为设定”且可以灵活调整,通常是为了使用上的便利性或特定读者群设计的。因此,即使分类方法存在差异,只要能够高效地服务于信息检索和管理,都可称之为“合适”的分类。
生物分类的特殊之处
但是,生物分类的本质却完全不同于图书馆里的“整理术”。虽然我们面对同样的信息爆炸——地球上已知生物约200万种,实际种数更可能高达千万乃至上亿种,如何分门别类、理清头绪无疑极具挑战性——但生物分类有一个图书馆分类所无法企及、极其特殊的属性:它理论上有且只有一个“正确答案”。
生物分类的标准不是人为便利、主观划分,而是客观、唯一的历史事实——进化谱系。每一种生物的位置,都映射着它与其他生物在亿万年进化树上的真实“亲缘关系”。
生命的演化史就是一幅巨大的家谱,每一个现生或已经灭绝的生物门类,都在其中拥有唯一的位置。我们之所以能用系统的方法给生物分类,并非因为我们喜欢把事物摆整齐,而是因为演化确实塑造了一个“真正存在”的生命谱系。这条进化路线如同人类家族的族谱,有明确的时间顺序和分支节点。分类学家的任务,就是寻找最科学的证据,将现存与历史上的生物准确地放回他们的进化“家谱”上。
因此,生物分类与图书分类在多个维度都截然不同。我们不妨通过下表进行对比:
正因如此,生物分类学既严谨又富有挑战,是揭示宇宙生命奥秘的重要科学。科学家们正不断完善分类体系,让它最大限度地接近进化历史的“真实”。我们每一次认识新物种、修正旧分类,都是在拨开演化树上的一层迷雾,向唯一正确答案更近一步。
传统分类的困惑与局限
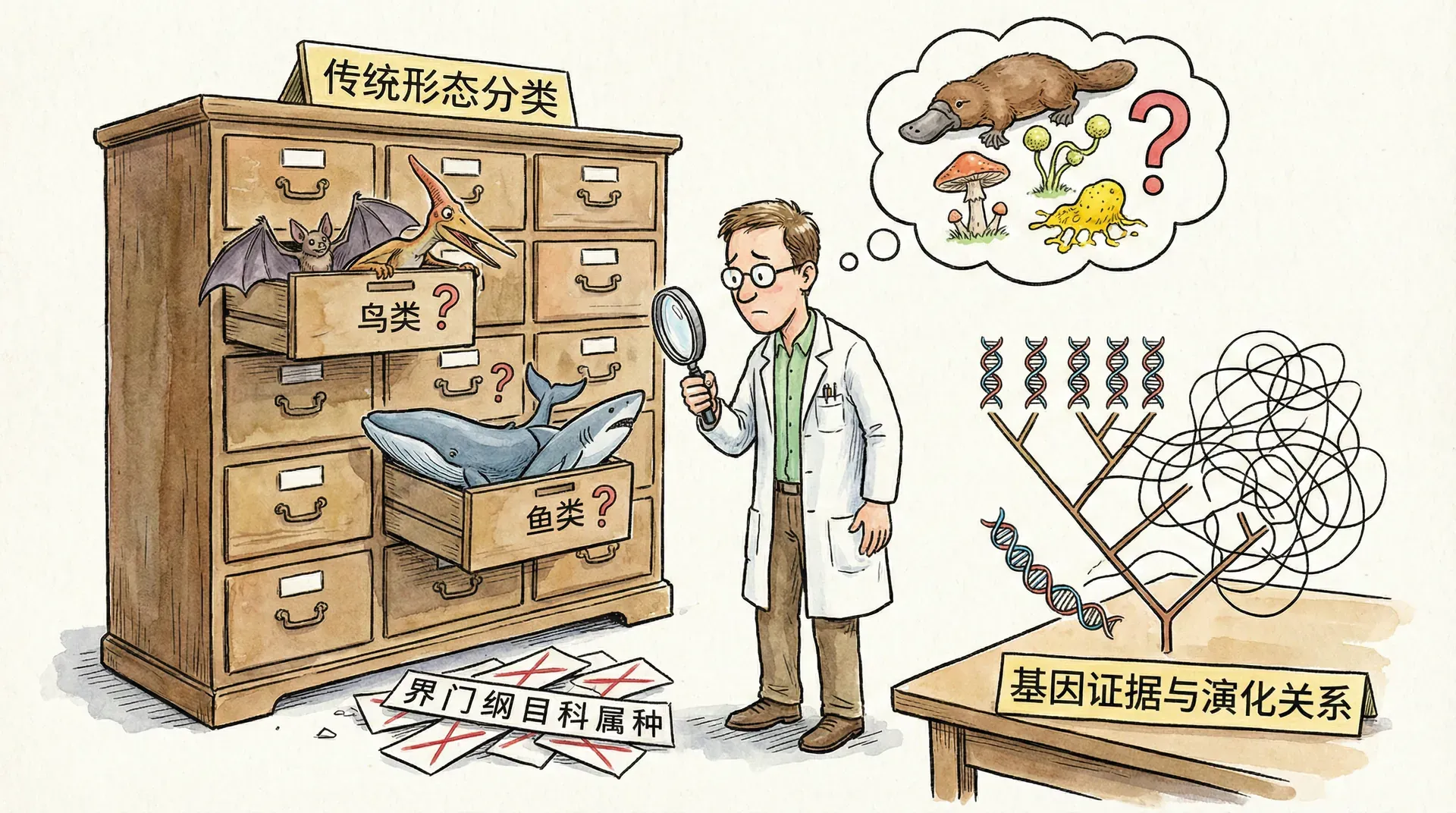
外观相似不等于血缘相近
早期的博物学家主要依靠生物的外部形态、颜色、体型等可见特征进行分类,这在自然观察和初级辨别中确实有一些实际效果,但随着生物种类和了解的深入,这种表面上的“相像”经常掩盖了真正的亲缘关系。你或许听过“相由心生”这句话,然而在生物世界,“外表决定内在”远远不是普遍规律——同样的外观背后,往往隐匿着截然不同的进化历史。
如果仅凭肉眼观察,有人会觉得华南虎和金钱豹极为相近——斑纹相似,体型都大,都是猫科猛兽、顶级捕食者。但分子生物学研究却揭示:华南虎和其他虎亚种(比如东北虎、孟加拉虎)才是“真正的一家人”,而离金钱豹的血缘其实更远,即便它们的斑纹在外观上看起来相似。
例如,中华白海豚和长江江豚都生活在中国的内海和河流中,有着流线型的身体以及极善游泳的特性。但从系统发育关系来看,白海豚属于齿鲸类,和江豚“八竿子打不着”,甚至在某些分支上,与河马的关系还更近于江豚。表面相似与真正的亲缘,往往不是一回事!
不仅如此,生活习性、生态环境的相似有时也会让完全无关的生物变得“似曾相识”。比如大家熟知的刺猬和豪猪,一个在中国分布广泛,一个更多见于热带和亚热带地区,它们全身布满硬刺,看似是“亲戚”,但实际上刺猬属于鼩形目,而豪猪的亲缘关系离得很远,本质上是两条“平行进化”的线路。
收敛进化
为什么物种会出现这种令人匪夷所思的表面相似?其根本原因在于收敛进化。指的是不同的生物为适应相似的环境或生态位,独立地演化出类似的结构、行为甚至生理功能。
我们可以类比两个毫无联系的工程师,在全球不同的地方、面对类似的问题时,或许会各自设计出几乎一致的解决方案。在生物世界中,这样的现象尤为突出。例如,“飞行”这一能力在人类历史上极具诱惑力,大自然中也不是只有一种动物掌握了飞行术。鸟类、蝙蝠和已经灭绝的翼龙都拥有翅膀,但三者的结构与来源大不相同:
尽管它们都能够飞行,但“翅膀”这一特征是在完全不同的祖先基础上,通过长期进化各自“独立研制”出来的。其实类似的例子还有鱼类、海豚、企鹅在水中游泳时表现出的流线型身躯和鳍形肢,是分别在鱼类、哺乳类和鸟类三大类群中独立进化出来的成果,并不代表他们关系接近。这些案例揭示了:收敛进化往往让我们在观察生物外观时产生误判,必须借助更深层的证据揭示真相。
大熊猫的分类争议历程
要说中国最具代表性的动物,非大熊猫莫属。这个“国宝级”的物种,也经历了一段充满争议和变迁的分类历程,成为生物分类学困惑与进步的缩影。
19世纪末,西方博物学者在四川山林首次“发现”大熊猫,第一眼看到它,几乎所有人都立刻被其“熊一样”的体型、黑白分明的毛色所吸引。但细看之下却又发现它拥有一些奇怪的地方,比如“伪拇指”(一块特殊腕骨,能帮助抓握竹子)的存在。如此特殊的特征让分类学家们“众说纷纭”:有的科学家主张它和熊是近亲,有些则认为它和小巧的浣熊同类,甚至有人把它单独列为一家!
大熊猫分类历史演变
这个历史告诉我们,外表的“特殊”和少数几个形态特征容易误导人们,只有深入遗传和分子层面,才能真正还原大熊猫的进化“家谱”。现代分子生物学最终揭示:大熊猫是熊家族的一支,与浣熊反倒没有近亲关系。
大熊猫的“伪拇指”其实不是手指,而是腕骨的一部分特化而成。它是对竹子食性的一个极致适应性进化,并不能作为亲缘关系的判断核心。例如,澳大利亚考拉(有袋类动物)也演化出“伪拇指”来抓树枝,但是它与大熊猫在遥远的进化树上才有共同祖先。
类似的分类争议在中外都有,例如穿山甲和食蚁兽的鳞片防护和专吃蚂蚁的习性;或沙漠蜥蜴、非洲变色龙的色彩变化、攀爬技巧。这些“功能收敛”都说明:单靠表象,无法准确判断血缘。
进化树
血缘关系胜过外在相似
那么,科学的生物分类究竟应基于什么?答案很明确:血缘关系。进化观念认为,所有的生命体都是通过连续不断的演化分支逐步产生的。每一次分化,都会出现一条新的分支,在进化树上留下清晰的节点。
如果将人类的家族关系画成树状图,你与兄弟姐妹之间的联系最近,因为父母是共同祖先。与堂、表兄弟姐妹间关系稍远,要追溯到祖父母、外祖父母。而更远的远亲,则需要往更早的世代追根溯源。生物系统分类也是一样:物种/属/科之间的远近完全取决于它们“最近共同祖先”分化出来的时间和顺序。
比如,华南虎和东北虎本质上几乎就是一个“大家庭”,大约万年以前才分开。而华南虎与金钱豹的共同祖先则要到几百万年前;而华南虎要和家猫“找到上一次亲戚聚会”,那已经是三、四千多万年前的古代猫科动物了。
支系分类学的核心思想
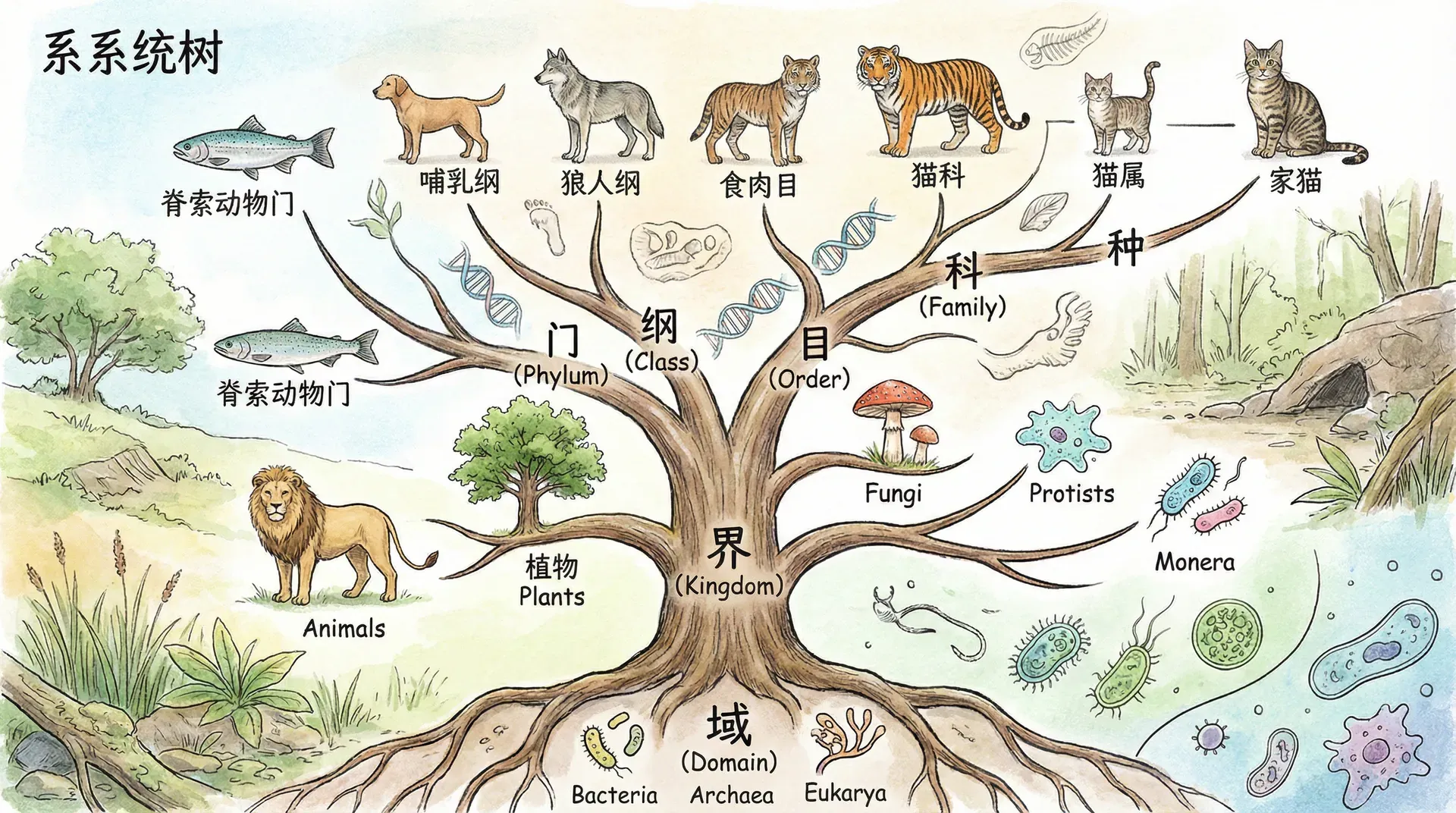
在20世纪中期,分类学发生了一场革命。系统发生学(支系分类学,cladistics)成为主导方法。它的根本思想很直白:只有拥有了最近共同祖先的所有后代组成的群体,才能被称为一个天然的分类群(单系群)。
而科学家们正是通过寻找不同生物的各种性状(外部形态、骨骼、行为、分子遗传等),逐步拼凑出谁和谁是一家、谁是“分出来的新枝”。进入分子时代后,DNA与蛋白质序列的比较成为“寻找血缘”的金标准,使进化树的重构更加精准。
以中国的珍稀动物为例,我们可以用一个“进化亲缘关系示意图”来直观展现这些物种彼此间的远近:
中国珍稀动物亲缘关系
现代生物分类正是要力求还原这样真正的进化树枝状结构,让每一种生物都“一锤定音”地归入自己的分支。
完美嵌套
最令人赞叹的是:生物分类体系有着任何人为分类都无法企及的“完美嵌套”结构。这意味着在真正根据亲缘关系划分的分类系统中,每一个更小的分类群都被完整包裹在对应的大群体内,绝对不会出现“重叠”、“混杂”或者“分家又合伙”的现象。
生物分类的嵌套结构示例
“完美嵌套”的秘密,就是每一个新的分支都是祖先的一部分,所有后代自动包含在更大的范围。分叉之后不再融合,这让生物分类变得异常清晰精准,能直观反映出生命进化的历史脉络。
如果我们不断向上追溯,每一个小分类嵌套在更大的层级之中,最终都指向所有生物的共同祖先。这是生物世界最宏伟的家谱,是其他任何分类系统所无法企及的。
分子证据
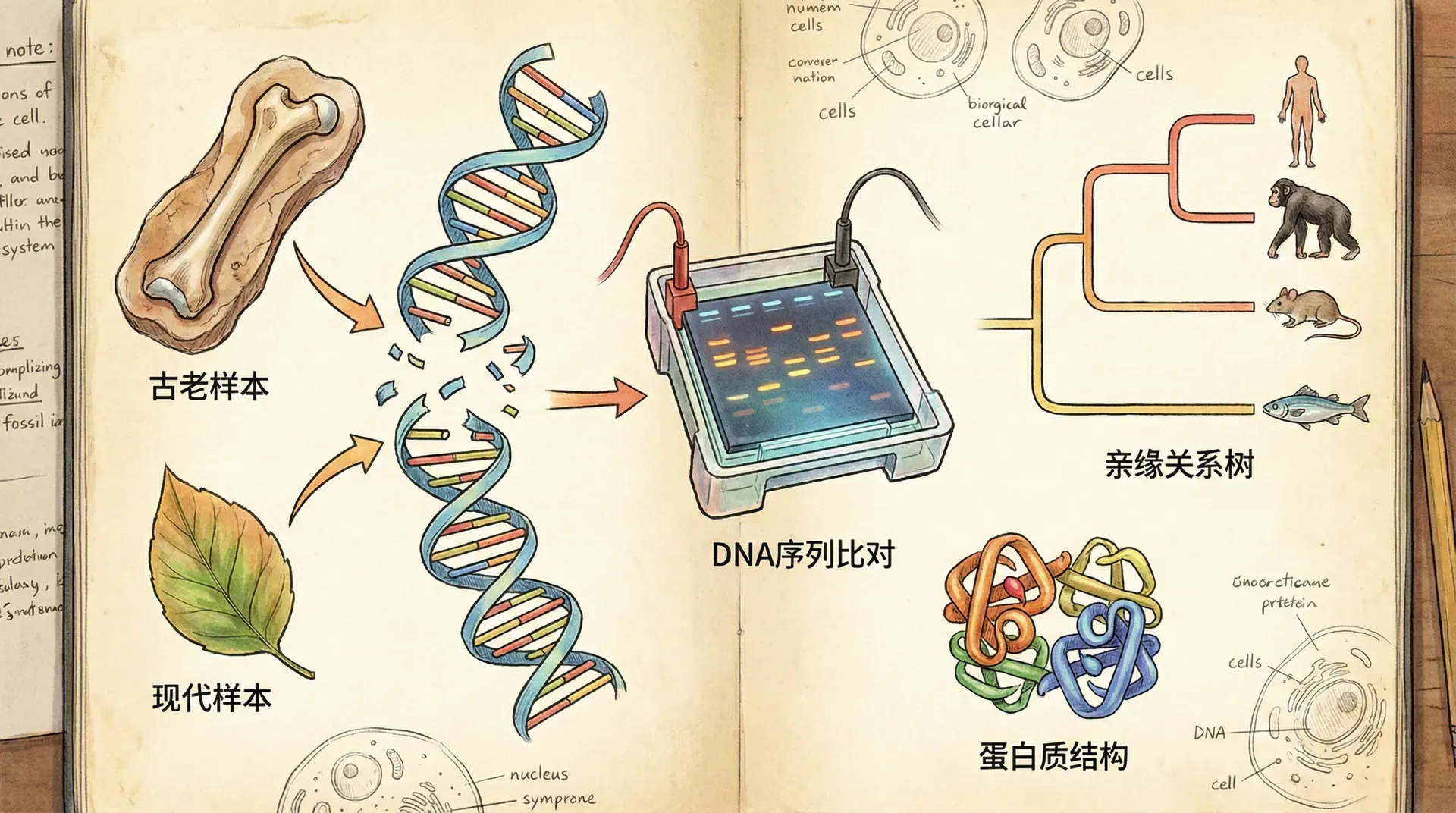
分子生物学的崛起,为生物分类带来了革命性的变化。通过分析DNA、RNA、蛋白质等生物大分子的结构与变异,科学家们获得了前所未有的工具来揭示生物间真实的亲缘关系,极大丰富并重塑了进化树。
生命的共同“语法”
20世纪下半叶,分子生物学的突破让科学家们发现:地球上所有生命——动物、植物、真菌、细菌,不论高等低等——都使用完全相同的遗传密码传递生命信息。DNA的四种碱基(A、T、C、G)组成“密码子”,其意义在几乎所有生物中都高度一致。例如,ATG无论在人、稻还是细菌体内,都是蛋白质的启动信号。这种“通用语言”意味着,所有生命极有可能拥有共同祖先,这也成为“共同祖先学说”最有力的分子证据。科学上把这一现象称为遗传密码的普遍性,为系统发育学和进化树奠定了坚实的理论基础。
正是遗传密码的普遍性,让科学家坚信:地球一切生命都根植于同一进化血脉。
分子时钟与进化历史
不同生物间DNA和蛋白质的序列差异会随着时间按相对稳定的速率积累。研究者依据“分子时钟”理论,对DNA差异进行比对、结合化石分化时间标定,便可推算物种分化的年代和亲缘远近。
以中国特有哺乳动物为例,测量大熊猫等动物的细胞色素C、血红蛋白基因差异,可以倒推它们分家的时间:
无论是细胞色素C还是血红蛋白,基因差异与分化时间几乎呈线性增长。由此,科学家重建出细致的进化树分叉节点,精度远超传统化石和形态学。此外,分子时钟技术广泛用于动物、植物、微生物甚至病毒等各大类群,揭示它们的演化路径与时间线。
DNA让“外貌”不再迷惑
分子证据同样避免了传统靠“形似”的误判。例如,大熊猫与浣熊都拥有“伪拇指”,外观习性相近,早期被视为近亲。但DNA比对显示,大熊猫与棕熊、北极熊的亲缘远高于与浣熊,证明“伪拇指”只是适应竹食性的收敛进化,不是直系血缘的证据。
这些数据表明,大熊猫与棕熊、北极熊实为近亲,同浣熊相距甚远。进化树脉络因此得以重绘。类似的分子纠偏还有:藏羚羊通过分子系统鉴定不属于牛科,而成独立支系;江豚等鲸类分类也因分子证据获得澄清。
上图显示,大熊猫与其他珍稀哺乳动物DNA相似度依次降低,反映不同物种间进化关系远近。这些分子数据不仅推动了分类修正,还为濒危物种保护与管理、科学繁育提供了坚实支撑。
现代分类学的方法与争议
分类学派的分歧与融合
分子分类技术带来的“证据浪潮”推动着分类学理论的不断进化,但在实际操作层面,不同学派之间的方法学争议却空前激烈。现代分类学并非“只有分子说了算”,关于“如何定义物种”、“如何构建进化树”的分歧依然存在。
主要分类学派对比
如今,多数分类研究实际上综合运用了上述多种理论和方法。科研人员往往在实际分类时并非拘泥于单一学派,而是融合形态解剖特征、化石证据、地理分布、行为生态,尤其是日益强大的分子生物学数据等多维证据链。通过对不同数据的权重与交叉验证,研究者能够得出更为可靠的分支关系与物种归属判断,使得分类体系既“接地气”又兼具科学严谨性和前沿性。这种多元融合方法成为当今系统分类学的主流趋势,也是未来持续提升分类准确性的重要保障。
分类准确性的不断提升
技术的进步,让分子分类在生物学中的地位逐步提升。过去,单一分子标记(如某一基因)只能揭示一部分真相;随着多标记、多基因组乃至全基因组测序技术普及,分类的分辨力几何级提升。
在不久的将来,全基因组测序有望成为分类学研究的“标配”,届时每个物种的进化位置都将获得空前清晰的画像。分子证据也将在医药开发、农业选育、生物资源保护等领域释放更大能量。分类学真正成为生命科学的“基石”,而不仅仅是命名与编目的工具。
随着分子技术的普及和不断创新,未来的分类科学无疑将更加智能、精准、客观,助力我们不断深化对地球生物多样性本质的理解和珍惜。
分类学与我们生活的联系
生物分类学并非脱离现实的学问,而是与我们的日常生活密切相关。了解生物之间的真实关系,有助于我们在实际问题中做出更科学的决策。比如,在生物保护、医药开发、农业生产等领域,正确的分类认识往往直接影响成败。
正确的分类认识直接关系到生物保护、医药开发、农业生产等多个领域的成败。
以医药开发为例,已知一种植物有药用成分,其近缘种往往也含有类似化合物。很多传统药材,如今通过分子分类学找到更多替代来源。在农业方面,分子分类学帮助识别作物野生近缘种,这些野生种为作物提供抗病、抗虫、耐逆等优良基因,促进杂交育种和基因改良。生态保护同样如此,只有准确厘清物种关系,才能制定科学的保护策略——同种不同群体应促进基因交流,不同物种则应防止杂交带来的基因污染。
分类学实际应用价值举例
生物分类学让我们认识到,地球生命构成了庞大复杂的家族树,人类只是其中一个分支。理解这种联系,不仅帮助我们深入认识生命世界,也提醒我们以更谦逊、负责任的态度对待其他生物。无论是高原雪豹、南海珊瑚,还是森林里的红松、龙血树,中国的每种生物都有独特进化与生态价值。科学分类研究不断揭示新的物种与联系,加深我们对生命本质的理解。
这正是分类学的魅力所在——它不仅整理和描述自然,更在探索生命奥秘、揭示自然规律。通过这门科学,我们不仅看清外部世界,也更好地理解了自身在生命网络中的位置。