反馈机制与演化理论
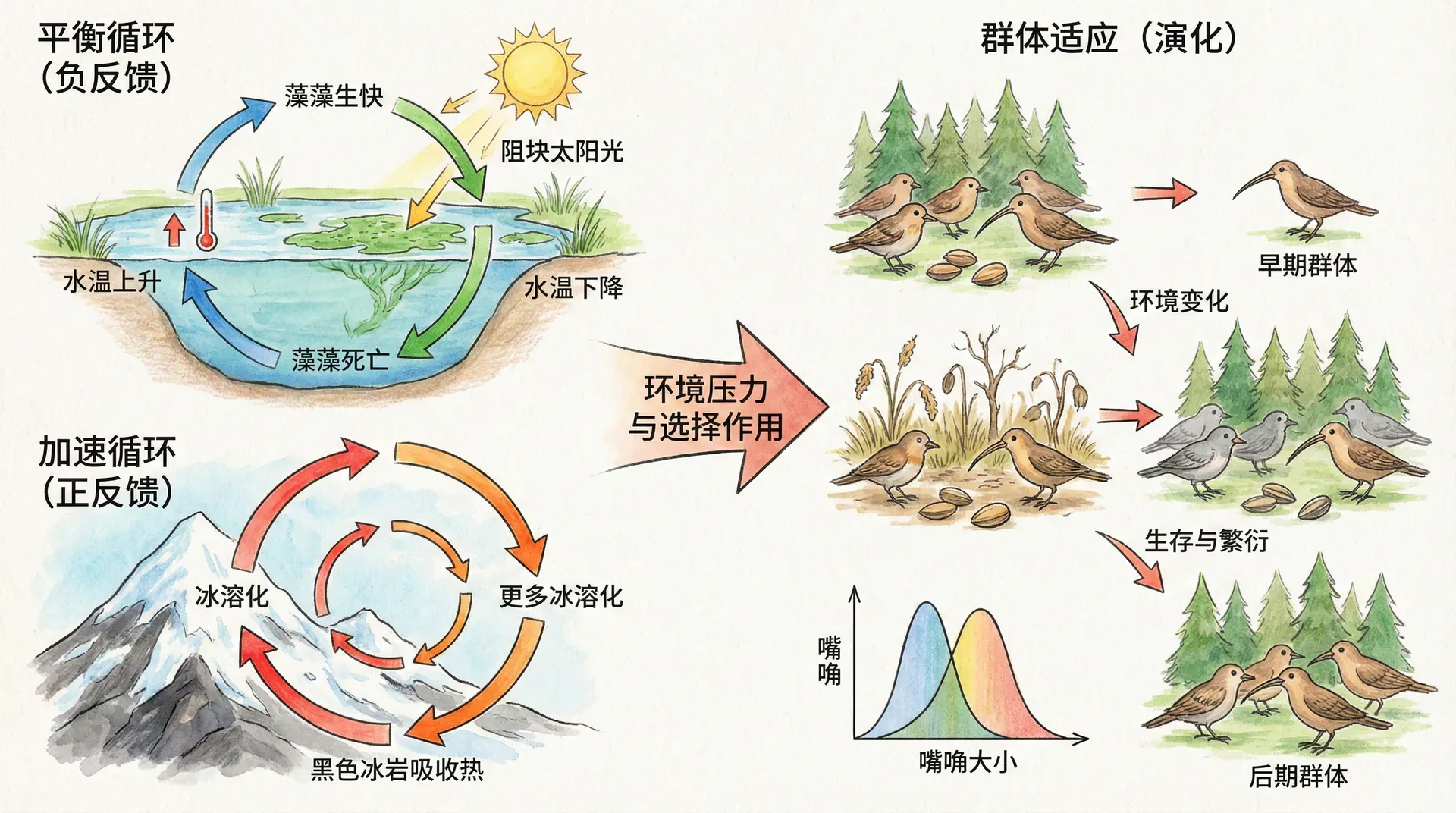
反馈机制是理解生物功能调节与复杂性演化的核心原理之一。在生物个体内部,负反馈帮助维持系统的稳定与平衡,例如体温、血糖等生理参数,通过持续调节应对环境变化,保障生命活动的有序进行。正反馈则在特定时刻引发系统性质的快速转变,促成生理过程的加速或完成,如分娩、神经信号的放大作用等。
更值得关注的是,反馈机制同样深度参与生物种群乃至生态系统层面的演化过程。演化理论指出,某些正反馈回路能够极速推动种群特性的累积变化,使得微小优势在选择压力下被放大,最终形成极端特征或复杂行为。这种正反馈强化的演化,不仅解释了物种适应性的提升,也揭示了自然界多样性与突变创新产生的动力来源。因此,无论在生理还是演化维度,反馈机制都为生物学搭建了调节、适应与演化的理论桥梁。
负反馈
负反馈是一种自我调节、自我抑制的调控方式。当系统的某项参数偏离理想范围时,负反馈机制会激活一系列调节活动,让参数“回头”,恢复到最适宜的平衡点。打个比方,就像家里的空调有一个温度传感器:当房间温度高于设定值,空调启动制冷;温度降到目标值,空调则自动停止或切换加热模式。这种方式保证了温度不会持续上升或下降太久。
在生物世界中,负反馈是几乎所有生命维持系统的基础。例如人体的体温调节,就是一个极其典型的负反馈系统。当环境温度上升,我们的身体通过出汗、皮肤血管扩张来散热降温;反过来,如果体温降低,身体就会通过肌肉颤抖产生热量,皮肤血管收缩减少散热。借助这种机制,人体能够实现大致恒定的体温(约37°C),无论外界如何变化。此外,血糖调节、血压调控、激素分泌等基础生理活动,几乎都离不开负反馈系统的保障。
负反馈机制是维持生物体内环境稳定的重要方式,包括血糖调节、血压调节、激素分泌调节、水盐平衡、免疫反应等多个复杂生理调控过程。它们确保生命活动的“过”与“不及”都能被自动校正,防止系统失控。
正反馈
与负反馈相对,正反馈则是一种自我增强、自我放大的反馈回路。当某个变化出现时,正反馈会进一步推动相关变化,使系统越来越偏离原始状态,直到外部机制或资源限制将过程终止。正反馈有时会造成“雪球效应”,初始的微小波动可能最终带来大幅度的结果。
实际生活中,森林火灾的蔓延就是一个常见的正反馈例子:起初可能只是地上一个不起眼的小火星,但当火烧起来后,产生的热量令附近的可燃物更易着火,火势随之变大——火越大,散发热越多,能点燃的范围也越广,最终形成非常可怕的连锁反应。如果没有外力(比如下雨、隔离带等)阻止,这个过程可以持续爆炸性增长。
在生物学领域,正反馈同样扮演着不可替代的角色。虽然总体上负反馈机制更常见、更基础,但一些关键的生命事件需要正反馈的“推动”——比如分娩过程。每当胎儿即将出生,子宫开始产生微弱收缩,这些收缩引起的刺激促使脑垂体分泌催产素,催产素又会增强子宫收缩,进一步加剧刺激,分泌更多催产素,形成正反馈环路,直到分娩完成。同样,在神经传导、血液凝固等过程中,正反馈也有着特殊功能。
通过上述分析,可以看到负反馈和正反馈虽然都是调控方式,却在生理现象和演化过程中产生了截然不同的结果。
正反馈在演化中的作用
除了在生理层面起到决定性作用,正反馈在宏观的生物演化过程中也展现了其“推波助澜”的独特威力。当我们观察到一些动植物拥有夸张且复杂的特征,比如雄孔雀那极其华丽的长尾屏、梅花鹿雄性的硕大鹿角、丹顶鹤引人注目的求偶舞蹈动作,往往会不禁疑惑:这些特征的发展似乎远远超出了单纯生存的需要。它们真的只是靠“随机变异和自然选择”逐步塑造的吗?
实际上,正反馈的演化机制可以带来“爆炸性”的变化。假如某种特征带来极小的优势,使个体获得更多繁殖机会,那么选择这类特征的雌性会越来越多,进而促进该特征进一步加强——正反馈由此启动。比如孔雀的长尾本来自普通,但是偏好长尾的雌性繁殖更多长尾雄性,长尾越来越常见且突出,最终演变为极为夸耀、几乎不利于生存的巨大尾屏。
上表展示了两类反馈机制在演化历程中的作用差异。负反馈控制下,生物特征大致稳定于“最适区间”,波动不大。而正反馈机制激活时,个别特征会呈现指数型爆发式增长,直至某一极限或资源、能量等外部因素制约其继续膨胀。
例如有些鸟类求偶色彩、鸣叫声调,或昆虫触角长度等都因为正反馈的作用而产生惊人的多样性和极端性。这些现象背后,常常隐藏着复杂的行为和遗传机制,如性选择、配偶竞争等。
这些令人惊异的演化特征,并不是单纯服务于环境适应,而更大程度上源于生物个体之间的相互作用(尤其是雌雄配对与选择)。当某一特征成为异性吸引力的标志时,它就进入了一个自我强化的快速演化轨道——假如没有外部制约,理论上特征可以不断“升级”,直到到达生理或生态的极限。
性选择理论
生物正反馈性状演化最经典的解释,来自达尔文的性选择理论。在19世纪末,达尔文在《物种起源》和《人类的由来》中提出了著名疑问:为什么雄孔雀会进化出如此硕大又累赘的尾屏?这种特征不仅绚丽夺目,还对生存带来负担,比如飞行笨重、更易被天敌发现,也需要消耗更多能量。达尔文认为,这些“累赘”其实是雌性择偶时青睐的性状标志。只要这样的特征能提升繁殖成功率(如带来更多交配机会、让基因更多传递),它即便降低个体生存能力,也会被不断放大——形成“越夸张→越受欢迎→再夸张”的自我强化正反馈。
性选择理论的核心观点是:即便某些性状会降低生存能力,只要能大幅提升繁殖成功率,就仍能被自然选择(尤其是性选择)保留。生存选择与性选择之间,始终存在着这样的“权衡”。
不过,达尔文的性选择学说最初并不被所有人接受。其中一个质疑是:为什么雌性会持续偏爱这些看似“无用”甚至连累生存的雄性?如果优先选择这些华丽特征,是否并会累积出更多“负担”让整个种群变弱?
对此,英国遗传学家费舍尔(R.A. Fisher)在20世纪30年代进一步深化理论,补充了性选择正反馈回路的遗传基础。他的解释揭示了性状极端演化背后的“跨代协同”正反馈机制,为我们认识生物多样性和极端特征爆发提供了关键钥匙。
费舍尔理论

费舍尔的理论提供了性选择极端特征正反馈演化的一个经典解释,这一理论既直观又精巧。为了更好地理解,我们以中国南方常见的红腹锦鸡为例,雄性红腹锦鸡拥有绚丽灿烂的羽毛和异常修长的尾羽,是自然界中最具代表性的“性装饰”之一。
假如我们把时光倒流到远古年代,当时红腹锦鸡的雄鸟尾羽长度只有如今的一半左右。自然界中基因存在变异,不同个体的尾羽会有长有短,而雌性红腹锦鸡天生对于配偶外观有着不同的偏好:有的雌鸟喜欢尾羽长一点的雄鸟,有的雌鸟则对此无所谓。这种“多样性”正是性选择机制产生动力的源泉。
尤其重要的是,参与调控雄鸟尾羽长度的基因,往往同样存在于雌鸟身上,而部分基因还会影响到雌性的择偶偏好。雌鸟自身虽然不表现“长尾”这一性状,但她们还是携带了和雄鸟一样影响羽毛发育的基因。从遗传的角度来说,这些基因在雌性体内仅仅不表达而已,却可以传递给下一代,无论是儿子还是女儿。
更有趣的是,负责“偏好长尾”的性状和“长尾本身”的性状可以通过遗传过程发生“联锁”效应,也即遗传学中所说的“连锁不平衡”。也就是说,一个雄鸟如果长有特别的尾羽,很大概率也继承了来自母亲的“偏好长尾”基因。所以他既长得漂亮,又有较高几率遇上偏好自己这种特征的雌鸟,从而繁殖优势极大提升。
这种现象的本质在于:“长尾”基因和尚且“偏好长尾”基因在同一家庭和同一谱系中累积。假如一只雌鸟喜欢长尾,她就倾向于选择长尾的雄鸟为配偶,她的儿子继承了长尾,而她的女儿则继承了“更偏好长尾”的倾向。因此,雄性的尾羽会一代比一代更长,而雌性的偏好也会一代比一代更极端,这两个性状相互伴随——从遗传学上来看,是“连锁”的。
这种“连锁不平衡”意味着两个相关的性状基因组合出现的频率大大高于完全随机的状态。这一机制使得“雄性夸张性装饰”和“雌性极端偏好”可以协同演化,推动特征走向极端甚至看似荒唐的程度。在现实世界中,这正是孔雀极巨大尾屏、鸟类华美羽饰甚至昆虫夸张触角背后的基因学根源。
自我强化的演化循环
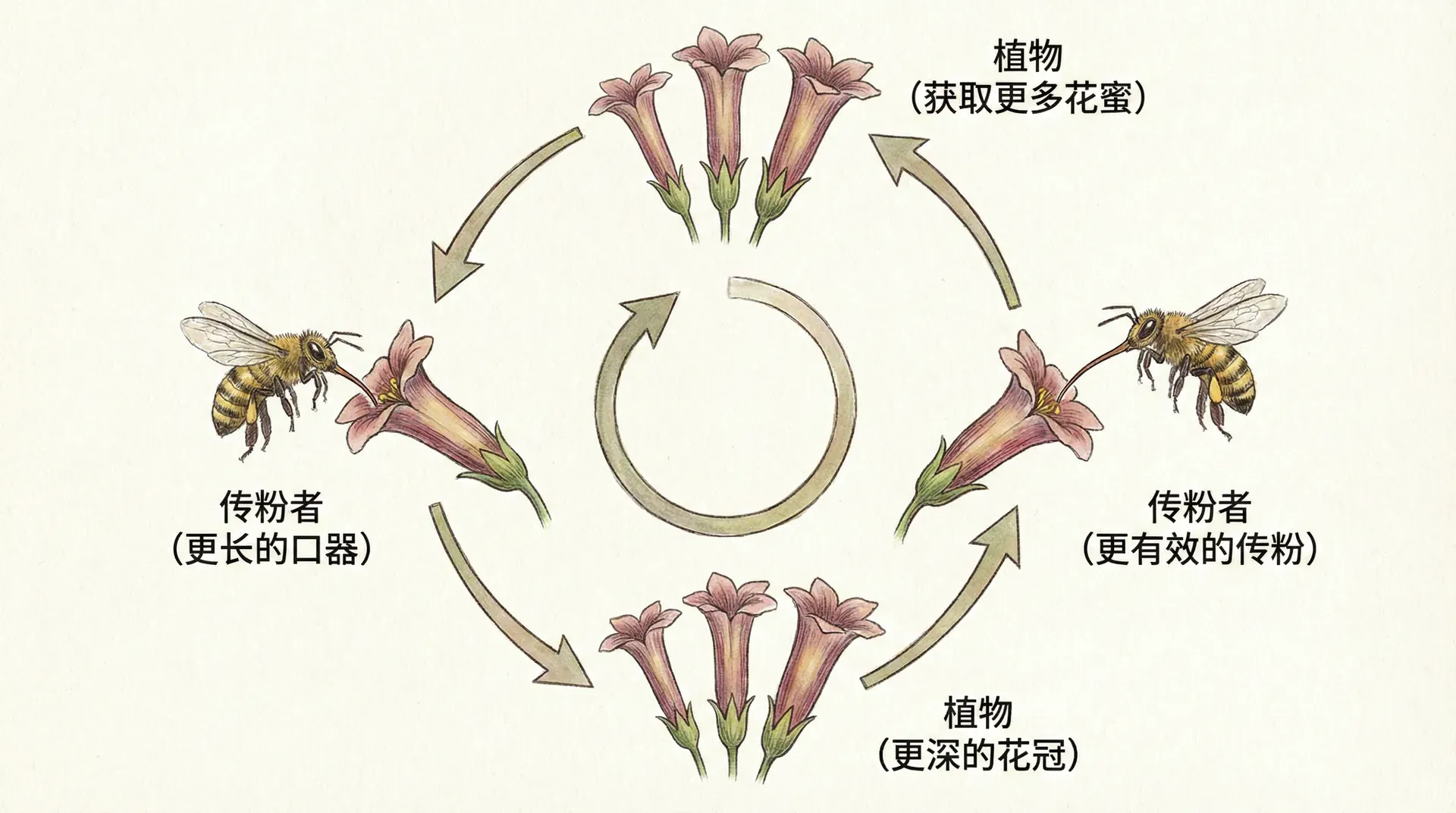
有了前面的基因基础,现在可以进一步理解为什么性选择中的正反馈极易“失控”。让我们回到红腹锦鸡的族群——假如大多数雌性都偏好尾羽更长的雄性,那么下一代会出现如下趋势:
- 雌性的儿子们更大概率长出修长尾羽;
- 雌性的女儿们更大概率喜欢长尾羽的雄性。
这个过程周而复始,每一代都会比上一代进一步“升级”极端特征和极端偏好。于是,一边是雄性尾羽越拉越长,另一边是雌性偏好日渐夸张,二者相互推动,像放大器一样没有明显上限——直至遇到生态或生理约束为止。
一旦“性装饰-偏好”联锁通路被激活,就会进入几乎难以逆转的演化快车道。每一次选择增强了优势基因的积累,也加强了下一代“择偶压力”和极端化趋势,从而成为正反馈的典型实例。
这种机制正是解释自然界中“性装饰爆炸性增长”的关键。例如红腹锦鸡闪耀的羽色、白冠长尾雉极端长尾、甚至甲虫和犀牛的巨大角,很多时候都源于雌性对特定性状的持续青睐,最终在遗传-偏好联锁的循环中,推动特征走向极端。
类似的循环可以发生在其他动物中:如有些鸟类的雄性复杂鸣叫、昆虫夸张的体型比例,甚至鱼类尾鳍的奇特形态。人类学家和进化生物学家也常用这一理论来讨论人类历史上某些“性吸引力”相关的行为、体态或装饰品的进化动力。
平衡与极限
虽然正反馈循环具有惊人的加速效应,但事实上这一过程不会没有止境地持续。正如森林大火终究会因燃料耗尽而熄灭,“性选择”的压倒性力量也不可避免地会被各种“实用性”压力所抵消。
以红腹锦鸡为例,过于夸张的尾羽虽然极具吸引力,但带来的弊端同样显著:
在性选择与自然选择拉锯的博弈下,最终族群中雄性的尾羽会稳定于某个“演化平衡点”——既能够最大限度增加吸引力,又不会致使生存成本失控,引发生态危机。
更广泛地看,正反馈机制推动性状“升级”,与负反馈机制(如天敌捕食、疾病、能量约束等)形成相对稳定的对抗与平衡。哪怕雌鸟的偏好再极端,实际生态系统的“约束力”仍然是决定特征极限的最终裁判。
现代研究的验证与实例
20世纪以来,大量实验研究和观察证据已强有力地支持了费舍尔理论及其预测。最著名的莫过于瑞典生物学家安德森(Malte Andersson)在肯尼亚开展的“寡妇鸟实验”。
寡妇鸟是非洲热带稀树草原常见的鸟类。雄性尾羽极为修长,远远超出生存所必需的程度。在安德森的著名实验中,他将捕捉到的雄性寡妇鸟分为三组:
- 实验组一:人工将雄性的尾羽剪短,制造“短尾组”。
- 实验组二:将剪下的尾羽用胶水黏接在其他雄鸟身后,形成“超长尾组”。
- 对照组:保持雄鸟原本的尾羽长度,未作处理。
在逐日的跟踪观测中,科学家们记录下各组雄鸟吸引来的雌鸟数量。结果表明:“超长尾组”雄鸟获得的配偶数约为“短尾组”雄鸟的四倍,甚至显著超过自然尾羽长度的“对照组”。这种跨越自然变异范围的“人为增强”,依然带来显著的“择偶优势”,说明雌性的偏好已经远远超出了当前雄鸟表型的平均水平。
安德森的实验不仅验证了雌鸟“超平均水平”的极端偏好,也支持了性选择正反馈理论的一个核心预言:只要偏好存在,夸张特征就有演化空间。这不仅适用于寡妇鸟,也适用于多个鸟类、鱼类、昆虫等动物——甚至在人类社会文化、审美、时尚流行中都可以找到相似的动力机制。
此外,其他实证研究也发现,在人工调控性状(如剪短或粘长鸟类、操作鱼类色彩、变更昆虫角的长度等)的实验中,只要雌性有明确偏好,相关性状就能在被选择中持续增强。反之,当生态环境变化,让夸张性状的生存成本快速上升时,这些特征也能在短时间内迅速萎缩。
综上所述,费舍尔理论和正反馈机制,让我们理解了为什么自然界充满了如此奇异、夸张、看似不利但却经久不衰的性装饰。它不仅为鸟类羽毛、昆虫角等提供解释,也为理解生物多样性的极端现象、人类行为演化、社会文化中“流行风潮”的本质,提供了兼具生物学与社会学意义的理论基础。
正反馈的其他生物学表现
除了性选择,正反馈在生物学中还有许多重要表现。这些现象帮助我们更全面地理解正反馈机制在生命过程中的广泛作用。
种群爆发现象
以东北地区周期性爆发的松毛虫为例。在常年条件下,松毛虫的数量受限于寄主树木、天敌、气温等多因素。但若遇连续暖冬、天敌数量降低或人为活动影响,种群生存条件改善,松毛虫数量就会开始攀升。
松毛虫种群增长过程中,正反馈循环表现为:
- 食物充足效应:最初密度较低时,每只幼虫都能获得充分食物,个体发育良好且成虫产卵量高,于是后代数量持续增加。
- 天敌稀释效应:当数量突破天敌的处理极限后,单位个体的天敌捕食概率下降,存活率进一步升高。
- 聚集优势:密集分布能提升越冬成活率、提高对寄主的定向定位能力,使整个种群对环境微小变化产生快速响应。
有时,一些人为因素也会干预正反馈,如林业病虫防治不及时、气候变暖助推越冬成功等,都会和上述机制叠加,导致几年的时间内虫口数量出现“井喷式”增长,从每亩几百只迅速爆发到几千甚至上万只,造成生态和经济的剧烈冲击。
这类生物种群的正反馈爆发不仅见于松毛虫,如蝗虫、鼠群、某些水生动物(如水母的“大爆发”)、藻类水华爆发等,背后都有类似的多重自我加强回路。
生态系统的级联正反馈与“相变”
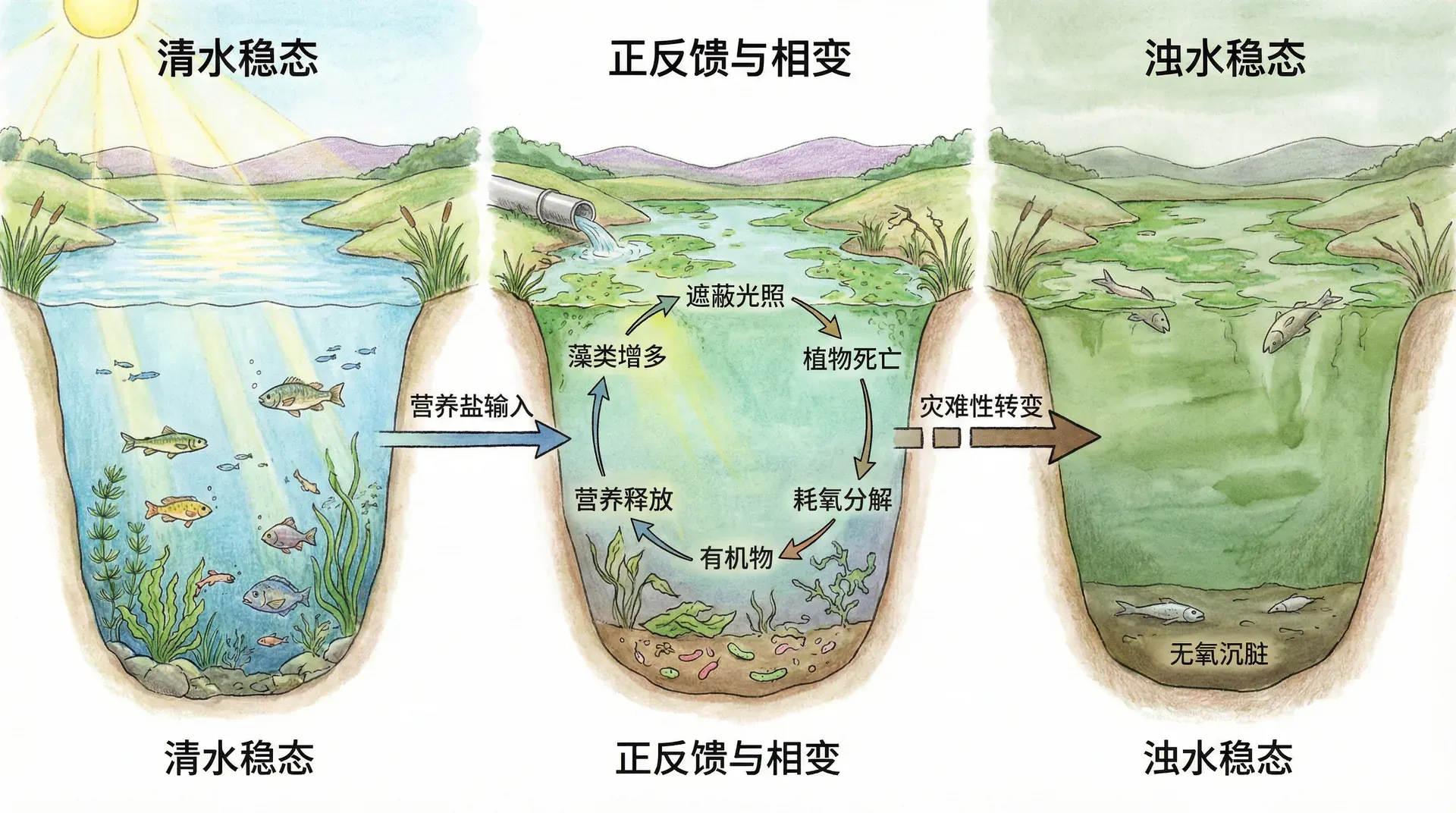
正反馈不仅局限于单一物种种群,在更大尺度的生态系统中,同样有“级联反应”。一个例子就是湖泊富营养化的灾难性转变。
当湖水中氮、磷等养分不断累积后,藻类大量繁殖——
- 藻类繁殖→覆盖水面遮光→水草死亡,失去稳态“过滤器”;
- 藻类死亡分解→消耗水体氧气→鱼类等高等水生动物因缺氧大量死亡;
- 死亡动植物分解→释出更多营养元素(如氮磷),进一步促进藻华;
- 水体流动性变差,微生物分解速度降低,导致“死水”循环;
这个过程中,系统的初始扰动被环环相扣地放大,最后湖泊生态系统实现了“从清水到死水”的灾变性跃迁(相变或者称临界转变)。国内如太湖、洱海、洞庭湖,国外如北美、北欧众多湖泊,莫不如此。
类似级联正反馈也见于森林火灾、草原退化、珊瑚礁白化等生态危机事件:一旦“锁链”上的任意环节正向加强,系统就可能加速滑向崩溃边缘。
生理系统中的正反馈
神经系统与生理调节机制也是正反馈经典舞台。前面已讲动作电位——神经细胞膜受刺激去极化,导致钠通道开放,进而电压越来越高,更多钠通道开放,形成猛烈的脉冲传导。这种机制保证了信号的“全或无特性”,使动物对刺激有极快反应与判别能力。
凝血过程也是正反馈的典型。人体受伤后,凝血因子被激活,每激活一个因子又能激活更多下游因子,级联反应令很少的初始信号迅速引发系统性变化,短时间形成血块止血。这使得我们能有效修补血管,防止失血,但过度凝血同样可能带来栓塞风险。
激素调控方面,排卵前雌激素升高,会刺激脑下垂体分泌大量促黄体生成素(LH),形成正反馈,最终促发排卵,同样是“极少信号-极大响应”的机制。
免疫细胞激活过程中,细胞因子释放吸引更多免疫细胞集结,炎症反应由微小病灶扩展至大范围——如果调控失控,甚至会产生“细胞因子风暴”等灾难性后果,这也是正反馈带来的负面效应之一。
文化现象中的类似机制
虽然文化与遗传进化路径原理有区别,但类似的自我强化模式在各种社会现象中屡见不鲜,尤其是在现代信息社会。
流行文化的病毒式扩散
抖音、快手等短视频平台上的“爆火”现象,就是典型的正反馈传播。最初一个内容被小范围用户点赞,算法据此加强推荐,扩大播放;更多人点赞/转发,则热度剧增,进入更强推荐池。每一轮反馈都让视频的受众和热度呈几何级数增长,形成“病毒式裂变”。
这种机制解释了为何少数作品可在短时间内席卷全网,让大多数内容始终“无人问津”。实际临界点在于何时正反馈足以压过自然流失(信息疲劳、审美疲劳等)的“负反馈”作用。
品牌,平台与社会认同的正反馈
在商业领域,苹果、特斯拉等品牌之所以能构筑坚不可摧的“护城河”,很大程度上依赖于正反馈带来的社会认同和口碑循环:
- 用户基数大→被视为潮流与品味的象征
- 社会认同感提高→更多人愿意跟风购买
- 用户体验与第三方服务更佳→正反馈驱动用户持续增长
这一循环并非产品卓越为唯一动因,多半依赖于“羊群效应”、主流媒体背书、社交网络传播等自我强化回路。类似逻辑还能拓展至社交媒体平台(如微信、Facebook的“网络效应”)、购物网站(淘宝、亚马逊“首选效应”)等现象。
学术界,体育,娱乐界的“超级明星”机制
学术界著名的“马太效应”可归结为正反馈的极致体现:少数高被引学者更易获得项目、合作者、媒体曝光,高水平资源进一步向这些“明星”学者聚集。这种机制同样适于体育、娱乐、网络红人领域——一旦实现流量“破圈”,资源便会高度集中,反而让“后浪”难以出头。
正反馈虽然推动个体向高处聚集,但也意味着系统整体多样性的下降、创新空间受限。因此,许多领域开始关注如何引入“反向激励”或“负反馈”机制,以避免过度集中、帮助新玩家突围。
总结
反馈机制是理解生物世界的重要工具。负反馈帮助生物维持稳定的内环境,而正反馈则能产生快速而剧烈的变化。在演化的长河中,正反馈通过性选择创造出了自然界最令人惊叹的美丽装饰。
费舍尔的理论揭示了这种“美丽”背后的深层机制:当基因连锁使得“吸引力特征”和“偏好基因”绑定在一起时,演化就会产生自我强化的正反馈循环。这个过程能够将最初微小的变异放大成极其夸张的特征,直到生存压力施加足够的制约。
从种群爆发到神经传导,从文化传播到品牌效应,正反馈机制在各个层面都发挥着重要作用。理解这些机制不仅能帮助我们欣赏自然界的多样性,也为我们思考其他复杂系统的演化提供了有价值的视角。
在这个充满复杂相互作用的世界中,掌握反馈机制的原理将帮助我们更好地理解生命现象的本质,也能为解决现实问题提供新的思路。