生物演化的建设性力量
生物演化常被认为只是一个淘汰不适应个体的过程,类似雕刻家凿去多余大理石,强调了自然选择对不利性状的“减法”作用。然而,这种观点过于简化了演化的本质。实际上,演化不只是消极地剔除错误,更是一股强大的创新与创造力量:在筛选个体的同时,不断组合、衍生和创新生物功能与结构,推动复杂性和巧妙设计的产生。
正是依靠这种持续的建设性力量,大自然才演化出丰富多彩的生命奇观。例如在中国,从高产水稻和多样竹林,到华南虎、大熊猫、梅花鹿及拟态昆虫等,都是演化创造力的见证。这些奇迹背后,离不开两个关键的建设性机制:一是基因间的高度协同合作,二是不同物种之间持续的竞争与相互促进。
演化的建设性力量主要体现在:其一,基因协作带来新功能和复杂性;其二,物种间竞争推动创新与适应提升。这两方面相互作用,共同推动了生物多样性和生命结构复杂化的进程。
基因协作机制与复杂性演化
基因间的协同适应现象
生物基因并不孤立地单打独斗,正如交响乐团的每位演奏家既要专注于自己的旋律,也需与全团严密配合才能奏出和谐的交响。细胞中的基因在表达和功能上,极其依赖于其“基因环境”——其它基因的表达状态、调控因子和信号相互作用。例如很多基因只有在某些细胞类型、特定的发育阶段才会启动,而且其功能效应受到主控调节基因的显著影响。比如,血红蛋白基因A和B只有在红细胞成熟过程中高效协同,才能保证氧气运输功能完整。
以水稻为例,一粒饱满的稻米形成,需要数十种与淀粉合成、细胞壁结构、抗逆性和养分积累相关的基因严密配合。例如:
- 淀粉长链合成基因:其表达高峰必须出现在籽粒灌浆的黄金期。
- 能量转运基因:确保为淀粉合成提供充足原料。
- 调控与辅助酶基因:调节上述基因的激活节律,决定灌浆速度和最终稻米品质。
如果其中任一基因表达“掉链子”,都可能导致稻米产量下降,甚至品质变差,抗逆力减弱。实际上,基因间的协同不仅表现为简单的“1+1>2”,而是在生物进化过程中经过反复整合和筛选,形成互补优势与复杂的正反馈网络。下表举例说明功能互补与基因依赖性的几种常见方式:
这种复杂协作,在高等植物、动物、甚至简单的细菌中都能找到踪迹。单一基因突变通常会被周围基因网络弥补,而长期不适应的变异则很快被淘汰——正是在这种“团队合作”下,生命复杂性和适应性逐步提升。
水稻淀粉合成系统的协作模式
以水稻的淀粉合成为例,这一过程就像一条高效协作的自动化生产线。每一类关键基因各司其职,并在特定时间点协同表达,确保稻米籽粒的充实饱满。具体来说:
- 淀粉合酶基因:负责将葡萄糖分子聚合成长链,为米粒构建能量储备。
- 支链淀粉酶基因:决定淀粉结构的分支度,影响米饭口感。
- 糖转运蛋白基因:输送原料到合成区域,保障能源供应不断档。
从上图可以看出,这些基因的表达高峰高度重叠,特别是在灌浆至成熟阶段(约60~80天)表现出同步性。这种“时间配合”是自然选择反复优化的结果,任何一个基因表达时序失调,都可能显著拖累水稻产量。
除此之外,水稻还包含大量调控性基因(如转录因子、感受信号通道等),在遇到干旱、高温或其它胁迫时动态调整主合成基因的活性,从而增加适应弹性。例如DREB转录因子在干旱激活淀粉合成相关基因,提升抗逆能力,这正是基因网络弹性的重要案例。
基因复制与功能分化
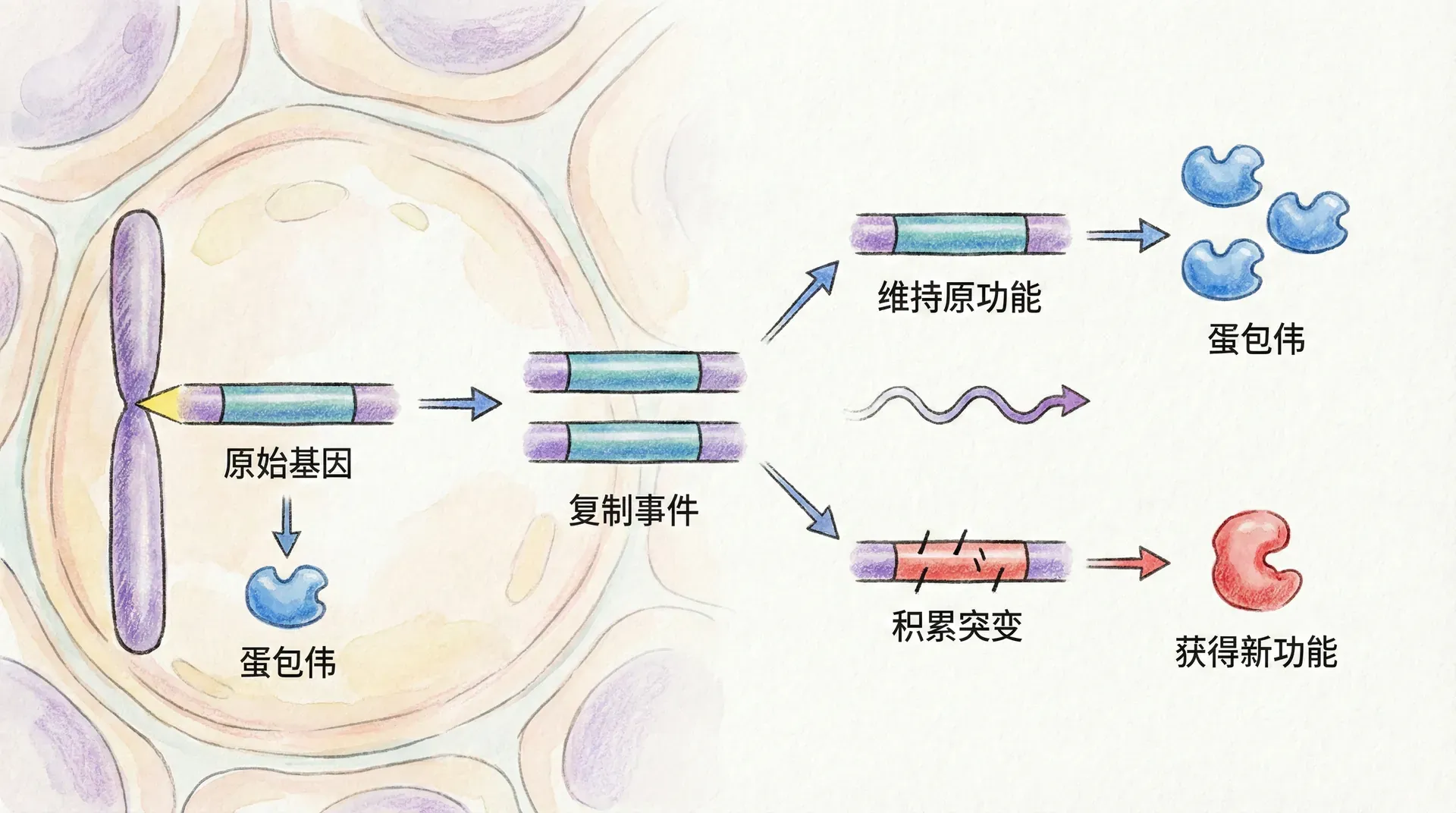
自然界并不会因为基因数量有限而止步不前。进化过程中,染色体复制、基因组加倍、片段重复等机制为生命提供了“冗余原料”。这些“同源拷贝”最初功能相同,但随着突变与时间推移,部分基因获得新特异化功能,或在不同环境下发挥差异化作用。比如:
- 一部分复制基因专精于耐盐碱性,另一部分偏向高光照条件下的效率调节。
- 有些复制基因用作基础生理功能,有些则成为环境压力下的“后备队员”。
这种分化大幅提升了生物的创新潜力和进化弹性。
再看农作物基因组的数据:
小麦因含三套染色体,基因总数居首。以抗病基因家族为例,常常出现“十几种专职岗”,每个基因分别针对不同病菌类型,形成强大的“分层防御”。高产作物往往正是依靠这种冗余多样的基因网,实现了产量、抗性和适应力兼备。
此外,基因复制还为物种创新提供了“试错空间”,即使某组基因出现突变损伤,完整功能仍可被同源基因补偿。
从单细胞到多细胞的演化历程
单细胞生命体的“全能作战”能力有限:一个细胞既要兼顾摄食、感知、繁殖,又受体积和功能极限束缚。多细胞生物则在协作和分工中迎来进化跃迁——部分细胞专责能量获取与代谢、部分专责信息传递、部分变为防御卫士,还有的进化为繁殖修复专能细胞。下表概述了单细胞与多细胞生命主要差异:
人体便是极致样例:由约10万亿个细胞组成,分工超过200种类型——如神经元传递信息、红细胞运输氧气、免疫细胞防御“外敌”。这幅总体协作的“生命城市”,每个细胞像不同职业群体互助共赢。
多细胞协作的机制同样出现在一些真菌、蓝藻和变形虫身上。例如,在环境恶劣时,粘菌的单个细胞能够快速聚集在一起,协同移动、寻找食物,最终集体变形形成孢子——这极大地展现了基因与细胞层面的灵活沟通能力。类似现象还可见于某些社会性细菌(如黄杆菌)的集群捕猎行为中。这些现象充分证明,生物演化的建设性力量远远超越了简单的“淘汰”,更体现在多层次的协作与新功能的诞生上。
生物军备竞赛与适应性演化
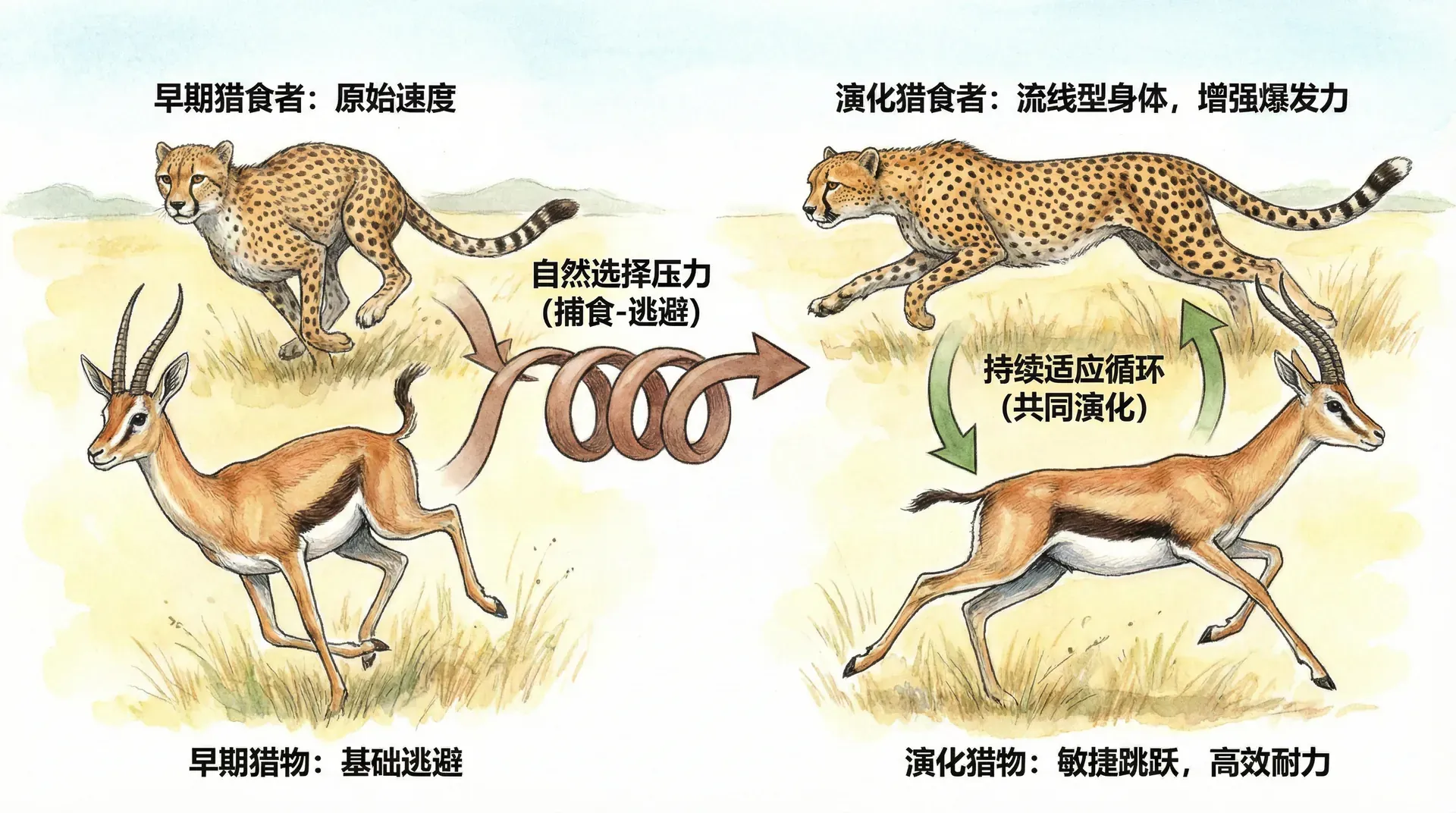
捕食者与被捕食者的演化博弈
如果说基因协作构成了演化机制内部的“内政”,那么物种间的军备竞赛则是激烈的“外交”。尤其是在捕食者与被捕食者之间,这种相互作用带来了持续的进化对抗——正如国家竞争,一方提升了攻击(捕食)能力,对手则必须不断强化防御,这个竞争周期便在自然选择推动下循环往复。
例如,在非洲大草原上,猎豹与瞪羚之间便上演了一场速度和敏捷竞赛:猎豹进化为体型更修长、奔跑更快、爆发力更强的顶级捕食者,而瞪羚则演化出极强的腿部肌肉和绝妙的急转能力,提高生存概率。它们之间的速度和敏捷竞赛,使两者的极限能力不断推升,但双方实际的“胜率”未见根本变化。
这种“双边升级却原地踏步”的现象,被称为红皇后效应。正如《爱丽丝梦游仙境》中红皇后所说:“在这里,你必须不停奔跑,才能保持在原地。”这描述了捕食者—被捕食者在进化压力下纵然都在“进步”,双方的竞争格局却保持动态平衡。
捕食与防御的演化策略
有些生物甚至会发展独特机制应对捕食压力,例如:
- 负鼠通过装死欺骗捕食者
- 刺猬依靠卷曲身体形成“铁刺球”
- 墨鱼不仅速度快,还能变色和喷墨云干扰视线
而被捕食者也会进化出群居、迷彩、剧毒等多种生存对策,体现出演化对抗的多样性和复杂性。
华南虎与梅花鹿的速度竞赛
在中国南方森林,华南虎和梅花鹿演绎着持久的“演化竞走”:华南虎不断选择出力量强大、敏捷且富有隐匿性的个体,而梅花鹿则逐代累积更敏锐的感官、高速奔跑和小群体生活方式,灵活应对捕食威胁。
观察数据可以发现:人类影响(如栖息地破碎)对两者种群带来急剧减少,过度干扰甚至打破了捕食者与被捕食者之间的微妙平衡。表面上双方的军备“停战”,但这会导致某一方适应性退化,生态系统整体稳定性下降。比如华南虎若绝迹,梅花鹿缺乏天敌,反而导致森林下层植被压力激增,生态链不再健康。
中外典型捕食—防御军备竞赛
对称与非对称军备竞赛
生物进化中的军备竞赛,常常表现为对称与非对称两种类型,各自有独特特点和代表案例。
对称军备竞赛
对称竞赛通常发生在同类生物间,主要是为争夺相同资源不断投入。例如竹林中,所有竹子都只能拼命长高以争取更多阳光,导致普遍高度增加、能量消耗剧增,但谁也没有获得“超额回报”,形成所谓“无效军备”。
另外,雄鹿间的鹿角大型化竞赛也是典型的对称军备竞赛:为赢得配偶,鹿角越来越大,最终代价远高于额外的繁殖优势。
非对称军备竞赛
非对称军备竞赛更常见于捕食——反捕食的攻防。例如蝙蝠发明了超声波定位,而夜蛾进化出可侦测和干扰超声的听觉与拟声。寄生蜂独特的产卵钻孔技术和某些蝴蝶拟态成毒虫等也属于这一类。非对称竞赛直接催生了生物在感知、外形、行为上的“策略创新”和多样性。
对称—非对称类型对比
红皇后效应
红皇后效应是演化军备竞赛中极具哲理的一环。它告诉我们:即便每个物种都在拼命提升适应能力,但由于对手同步进步,双方竞争地位并不会发生质变。
例如,蝙蝠与夜蛾之间持续上演“声纳大战”:蝙蝠进化出精细的回声定位系统,夜蛾则开发出能够感知并干扰声波的耳朵和反制技巧。随着进步加剧,双方能力整体显著提升——但“成功率”却并无质变。这说明了军备竞赛在推高技术门槛、累积整体适应潜力的同时,并不一定带来个人或某一代的压倒性胜利。
红皇后效应不仅存在于动物之间,也广泛表现于其他领域,如:
- 病毒与宿主间的“免疫—入侵”竞赛(如新冠病毒不断变异,人类免疫系统持续优化)
- 人类与细菌间抗生素—耐药性之争
- 植物与害虫/病原之间的化学武装竞赛
红皇后效应本质上是适应性演化的动力源泉——不进则退、进步即维持。这正是自然界多样性和创新性的深层机制之一。
演化的经济学原理与平衡机制
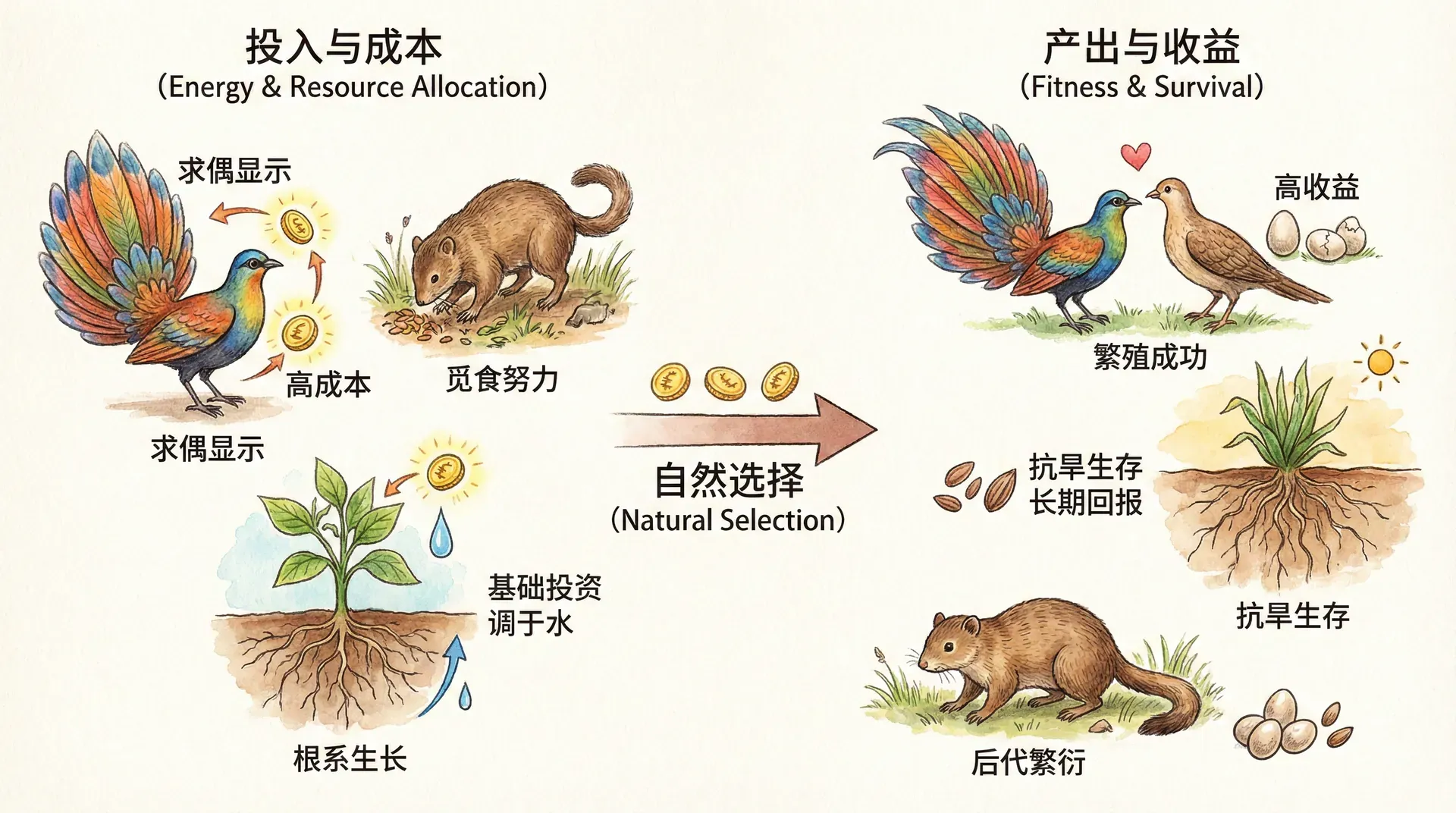
成本——收益、囚徒困境与进化均衡
生物的演化并非“更快、更高、更强”地无止境升级,每一步改进都需权衡成本与收益。从自然选择视角看,进化是一场有限资源下的分配博弈:鹿想跑得更快,必然要把更多能量投入肌肉生长;竹子想长得更高,也需追加资源于支撑结构和能量转化。这意味着任何对某项特征的“投资”,都会带来对生存、免疫、繁殖等其他领域的机会成本。
合理的资源分配直接影响种群未来——分配效率更高的个体,其基因能更好流传后代。反之,某些性状如果发展过度(如鹿角极度巨大化),一旦成本超过收益,反倒会因负担过重遭到淘汰,整个种群最终会达到“适度均衡”。
这种资源竞争和均衡的经典案例正是竹林高度竞赛:如果每棵竹子都能“协议”长到合适高度,就能最优吸收阳光、节省资源,利于繁殖和抵抗病害。但囚徒困境注定了,只要有一棵竹子“偷偷长高”,其它竹子也不得不跟进,最终大家都消耗了更多资源,却没有谁真正获益。类似现象在人类社会、经济和教育中也屡见不鲜,比如“分数大战”、企业广告投入等,最终形成集体次优局面。
军备竞赛的终止条件
生物军备竞赛虽然不断推动新特征演化,但也不会无限制升级。其终止或减缓受两方面约束:
- 物理极限:如奔跑速度受骨骼与关节极限所限,植物高度受制于茎秆结构和风压。超过极限将导致结构失稳,投资无回报。
- 经济约束:投资的边际回报会递减,甚至为负。例如动物为求更快,心肺器官过度发达反碍其他生理机能。最终,自然选择“调优”所有特征,使其收益与代价达成平衡。这也解释了为何现实中没有“无敌生物”,一切性状都是多重妥协下的最优结果。
演化的经济约束原理说明,所有生物特征均是各种选择压力下达成的最优平衡,在特定环境中体现出解题智慧。
中国生态系统的“天然实验室”
中国复杂多样的自然地貌(海洋、平原、森林、高原、沙漠等)为进化博弈和适应性创新提供了丰富“样本”:
- 长江流域:中华鲟、江豚等顶级捕食者与鱼类、水生昆虫之间展开层层军备竞赛。昆虫发展出伪装或透明化,鱼类进化感知与速度,江豚则拥有高频声呐感知,攻防之间基因网络持续优化。
- 青藏高原:极端气候塑造了独特适应体系。藏羚羊耐缺氧血红蛋白、超强耐力,能高速躲避狼群。高原特有植物如垫状草、龙胆等通过矮化、色素沉积降低能量消耗及抵抗恶劣气候。这一切都在物理极限和环境约束下达成精妙平衡——狼与藏羚羊之间的军备竞赛,最终也要服从极端高寒的“第三裁判”。
随着城市化发展,人类活动正在重塑新的生态较量。例如城市中麻雀、喜鹊等鸟类与天敌(如猫、隼)之间通过警觉、逃逸及新型栖息地选择展开持续博弈,新型军备竞赛不断出现。
从竹林的囚徒困境,到高原的极限适应,再到城市生态的新军备,生物进化史就是一场协作与对抗、创新与约束的“动态均衡”演化。基因协作提升内部系统效率,军备博弈则激励跨物种创新。理解这一切,不仅能更敬畏自然、洞悉生态微妙,也为农业、疾病防控、生态保护等人类实践提供深刻启示。毕竟,无论自然还是社会,只有在竞争与协作并重、理性权衡成本收益的条件下,才能实现持续的最优发展。