生态系统的能量流动
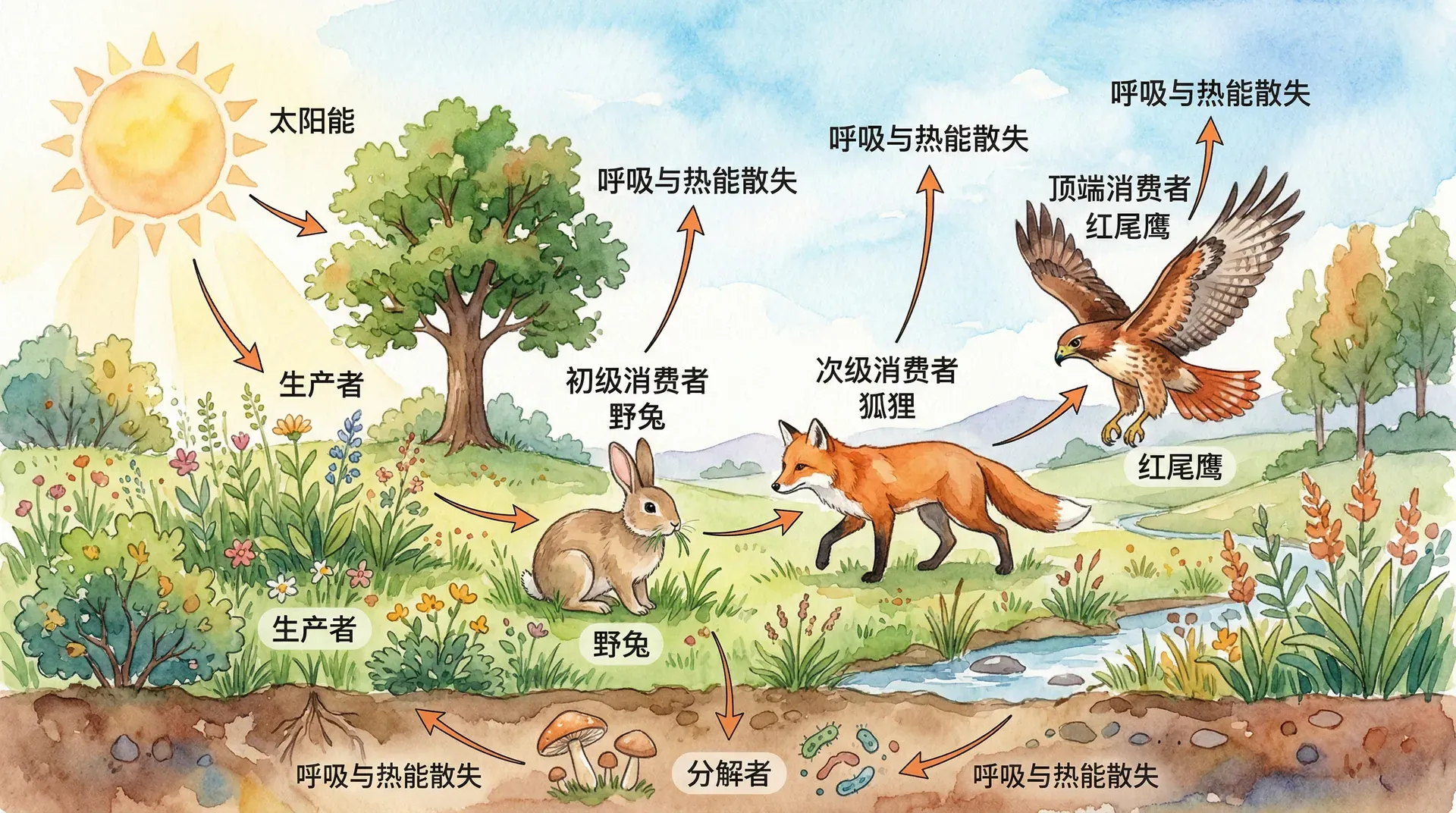
能量是生态系统运作的基础动力。无论是阳光透过云层洒在森林树冠,还是风吹动草原上的微小生物,能量在每一个生态系统中都是不可或缺的驱动力。在生态系统内部,能量经历着一系列复杂的流动和转化过程:白天的第一缕阳光被绿色植物,通过光合作用固定下来,转化为有机物中的化学能,为整个生态系统的生命活动提供“燃料”;这一能量随后被初级消费者(如植食性动物)获取,再逐级传递到次级、三级消费者(如肉食性动物),直至顶级捕食者。而当动植物死亡后,分解者(如细菌和真菌)又将有机物分解为无机物,将部分能量以热能形式散失到环境中,完成了能量流动的闭环。
理解能量在生态系统中的流动,有助于我们从整体上把握自然界的规律。例如,为什么自然界中生产者(如植物)的数量远多于消费者?为什么食物链不能无限延长?为什么保护顶级捕食者还必须要保护其下游的整个生态系统?这些问题背后,都是能量流动与转换的基本原理在起作用。
能量流动不仅是生态学研究的核心,也是生态保护和资源管理的重要理论基础。只有深入理解能量如何流动和耗散,我们才能科学地评估不同生态系统的生产力、制定有效的保护措施,以及合理利用自然资源,实现人与自然的和谐共生。
能量的来源与生态系统的驱动力
生态系统中的能量绝大部分主要来源于太阳辐射。太阳不断释放巨大的能量,日复一日,地球表面每天接收到的太阳能约为1.73×10¹⁷瓦,但实际上,能够被绿色植物通过光合作用固定下来的太阳能量仅仅占总量的1%左右。这个比例看似微小,实际上却为地球上的生命活动提供了源源不断的能量保障。没有这1%的能量输入,地球上的生命体系将难以维系。
在我国的长白山森林生态系统中,科学家通过多年监测数据发现,每年每平方米森林可获得的太阳辐射能量约为5×10⁹焦耳,而其中只有约5×10⁷焦耳(约1%)被植物通过光合作用固定并储存为有机物中的化学能。这部分被植物固定下来的化学能量不仅供给了植物自身的生长和呼吸,还通过植食性动物、肉食性动物等不同营养级逐级传递,最终构筑起以昆虫、鸟类、中小型哺乳动物,直至东北虎等顶级捕食者为代表的复杂生物共同体。这一切,都是建立在最初被固定下来的太阳能量基础上的。
植物通过光合作用将太阳能转化为化学能,这是生态系统中几乎唯一持续而有效的能量输入途径。虽然光合作用的能量转化效率相对较低,只有1-2%,但太阳能输入总量巨大,因此依然可以支持地球上复杂且多样的生物圈。这也说明了植物在维系全球能量平衡中的核心作用。
在一些特殊的环境中,由于地理和气候条件的限制,能量的输入和利用效率会显著不同。例如,在青藏高原的高寒草甸生态系统,由于这里的海拔高、气温低、日照但生长季节极短,植物的光合作用强度和效率均受到限制。研究数据显示,该地区的植物每年固定的太阳能约为温带森林的40-60%,远低于低海拔地区。
但即便如此,有限的能量输入依然支撑着如藏羚羊、野牦牛等具有高原适应性的典型物种生存和繁衍,展现了生态系统对外部环境条件的惊人适应性和生物多样性的独特价值。与此同时,在沙漠等干旱生态系统内,能量输入的波动性更大,植物和动物也需要通过各种适应机制(如储水、昼夜节律变化等)来缓解能量与水资源的时空不均衡。
与此同时,少部分生态系统(如深海热泉)可以通过化能合成作用获得能量输入。例如,某些化能自养细菌能够利用地壳释放出的化学物质作为能量源参与能量转化,从而构成特殊的生态系统。但就地球总体而言,绝大多数生态系统依赖太阳能驱动。
食物链
食物链是生态系统能量流动的核心线索,它描述了生物之间因“吃与被吃”而发生的能量转移路径。在最理想化、简单的情况下,一条典型食物链包括生产者(如绿色植物)、初级消费者(如植食动物)、次级消费者(如肉食动物)等基本环节。但实际上,食物链往往包含更多层级,结构也更为复杂。
以我国东北森林草原交错带为例,如下信息展示典型的陆地食物链结构:
而在太湖水域的水生生态系统中,能量流动如下所示:浮游植物(生产者)通过光合作用吸收太阳能,浮游动物(初级消费者)取食浮游植物,小型鱼类(次级消费者)以浮游动物为食,大型肉食性鱼类(三级消费者)捕食小鱼,整个能量沿营养级递减传递,形成完整链条。
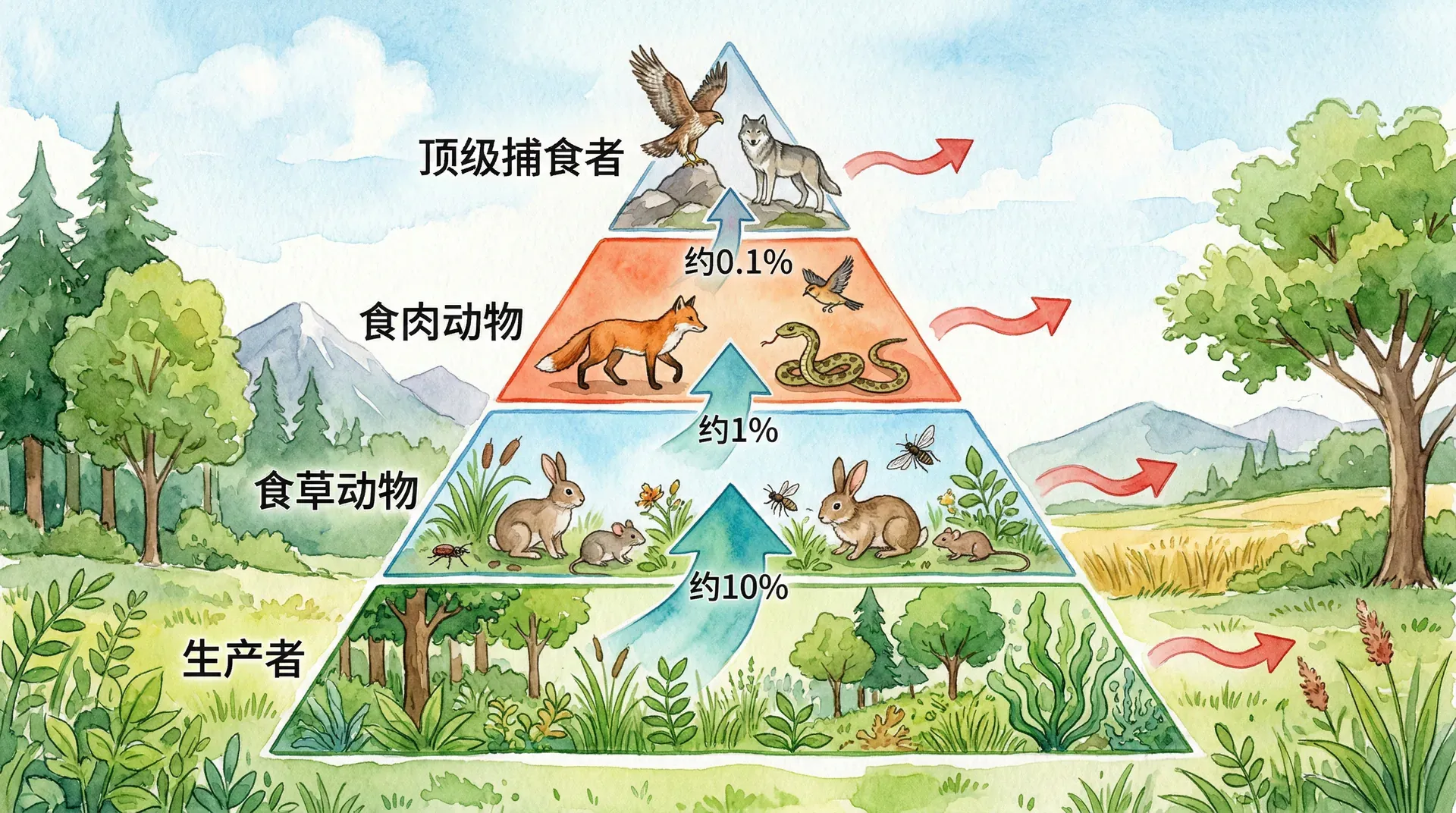
上图显示能量每通过一个营养级,就会大幅递减,一般只有10%左右能量能传递给下一级,其余90%在呼吸代谢、热量散逸等过程中损失。因此,生态系统内的能量呈现典型的“金字塔”分布,这一规律限制了食物链的最大长度,也说明了顶级捕食者稀少的根本原因。
在自然生态中,真实能量流动往往不是线性的单条链路,而是交错复杂的“食物网”。举例来说,在长江中下游湿地,芦苇不仅能被昆虫取食,还是多种鸟类栖息觅食场所。昆虫既为蜘蛛、青蛙、小型鸟类提供食物,这些动物又可成为蛇、猛禽等更高级捕食者的猎物。正因如此,生态系统面对物种减少或环境扰动时,更具弹性和自我调节能力。
另外,生态系统中还包含“杂食性”“腐食性”动物。例如,棕熊既吃浆果也捕食鱼类;分解者(如细菌、真菌)则分解有机遗体,将能量带回循环。综合来看,生产者固定太阳能,能量沿营养级递减并交错流动、形成食物网,是生态系统运行和稳定的基础。
营养级与能量传递效率
营养级用来描述生物在食物链中的功能位置。第一营养级为生产者(绿色植物、藻类等),依靠光合作用为生态系统输入初级能量。第二营养级为植食动物(初级消费者),直接依赖生产者。第三营养级为肉食动物(次级消费者),以初级消费者为食。部分生态系统还有杂食动物与分解者,使营养级结构更具弹性。
不同营养级之间的能量传递效率——通常称为“林德曼效率”——平均约为10%。下表综合展示了营养级、能量来源与典型传递效率:
造成能量损失的原因主要有三:
- 前一营养级部分生物量难以直接消化吸收,如植物纤维或木质部;
- 动物摄入的食物只有部分被消化,剩余以粪便排出体系;
- 真正被吸收的能量又大部分用于生命活动(如新陈代谢、运动、繁殖),以热能形式散失。
如上图和数据所示,能量传递效率在不同营养级、不同生态系统和取样时间中略有波动,但整体规律一致,呈现逐级递减。比如生产者到植食动物约为8-12%,肉食动物之间则更高达15-18%,原因在于动物性食物消化率更高。
以“中国东北农田‘玉米-蝗虫-青蛙-蛇’”为例,每公顷玉米每年可固定2×10⁸千焦耳,蝗虫摄取约2×10⁷千焦耳(10%),青蛙获取约2×10⁶千焦耳(10%),蛇最终能量约为3×10⁵千焦耳(15%)。如果病虫害压力增大、农田结构失衡,整个链条的能量效率会进一步降低。
全球各类生态系统(如极地苔原、热带雨林、草原、淡水与海洋)都存在类似结构。能量递减的“金字塔”分布不仅塑造了种群分布和数量,也决定了生态系统结构与稳态。
正是因为能量传递效率低,大多数生态系统的食物链长度通常不会超过4~5个营养级。再向上传递,已无足够能量支持更多或更大型生物的持续繁衍和维持。这一规律对生物多样性、资源利用和生态保护有重要启示。
生态金字塔的三种类型
生态金字塔是用图形和数据表达不同营养级关系的常用方式,分为数量金字塔、生物量金字塔和能量金字塔三类。
数量金字塔:表示各营养级生物个体数量。在普通的陆地草原系统中,通常最底层为大量草本植物,上面是数量较少的食草动物,顶部是稀少的肉食动物。以华北平原落叶阔叶林为例,结构如下:
当生产者个体巨大时,金字塔可能“倒置”,如一棵大树可承载上万昆虫。
生物量金字塔则展示各营养级的总生物量(干重),更能反映能量转化状况。以内蒙古草原为例,每公顷草地干重植物4000千克,植食动物400千克,肉食动物约40千克,量级递减明显。
能量金字塔以各营养级能量流动速率或储存量为依据,始终为正金字塔型,因为能量顺热力学定律沿营养级下降。
图中可见:能量金字塔的递减更为剧烈,因为它代表能量流动和损失的动态过程;而生物量金字塔反映生态系统某一时刻的生物量积累,递减相对缓和。
如在典型的海洋系统,浮游植物的生物量虽小,但因繁殖极快,全年能量产出却大大超过摄食它们的浮游动物。这种情形下,能量金字塔仍为正金字塔型,而生物量金字塔可偶现“倒置”结构,进一步显示生态系统结构的多样性与适应性。
能量流动的定量分析
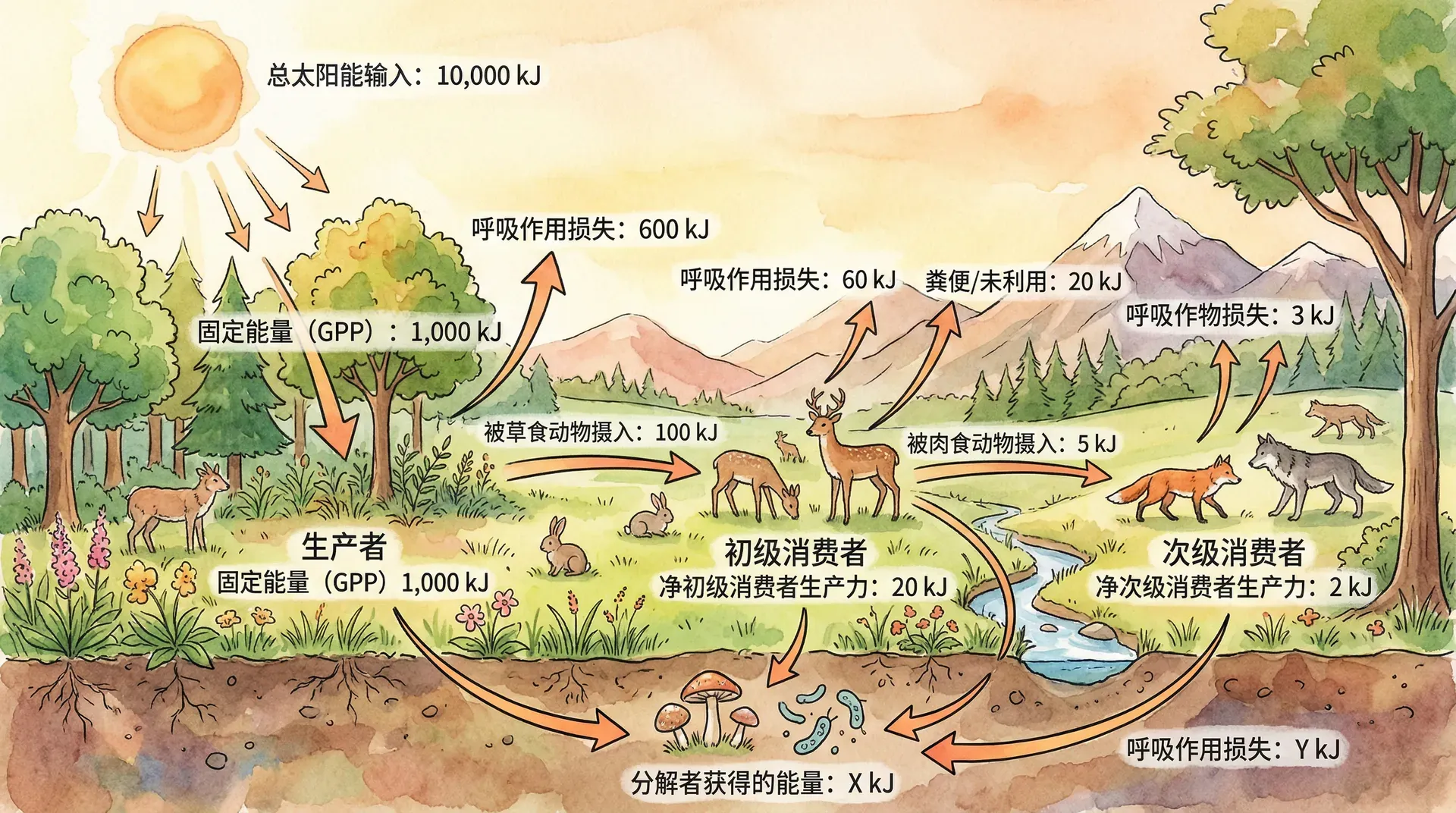
对生态系统能量流动的定量分析,是理解和管理自然与人为生态系统的重要基础。通过定量研究,生态学家可以揭示不同营养级间能量分配的比例、流动过程与效率,以及影响因素等,指导生态工程实践与资源可持续利用。
生态系统能量流动一般遵循如下路径:生产者(如绿色植物)利用光合作用将太阳能转化为化学能,产生“总初级生产力”(GPP)。生产者呼吸时消耗部分能量(R),GPP 扣除 R 后即为“净初级生产力”(NPP),可供消费者与分解者利用。动物(消费者)通过摄食获得能量(摄入量 I),部分经过消化吸收形成同化量(A),同化量中有部分用于自身呼吸,其余则转化为生长和繁殖的“生产量”(P),并成为更高营养级的能量来源。分解者则将有机废弃物和尸体中的能量部分释放,剩余以热的形式散失。
常见参数及意义如下:
我们以洞庭湖湿地芦苇沼泽为例说明能量流动链条:芦苇每年每平方米固定太阳能(GPP)约45000千焦,呼吸消耗18000千焦,剩余27000千焦成为NPP。植食性动物仅可摄入8000千焦,其中4000千焦被成功消化吸收(A),其它随废弃物排出。再扣除动物呼吸消耗约2500千焦,约1500千焦用于动物生长及繁殖,被更高营养级利用。这显示了能量逐级递减及能量流动各环节的定量关系。
能量流动过程还受外界环境显著影响。以柱状图呈现芦苇沼泽生态系统全年能量输入与传递动态:
夏季(6-8月)能量流动明显高于冬季,这与植被生长状况和气候密切相关。这种分析同样可应用于其他生态系统的生产力与承载力评估。
能量流动定量分析不仅用于自然生态系统,还指导农业等人工系统的优化。例如江苏稻田生态系统中,采用科学管理可将水稻产量的能量转化效率由30%提升到60%以上。同时,能量流动研究有助于生态系统可持续性评价和服务功能的合理配置。
随着遥感、温室气体自动分析等新技术发展,能量流动的精细化、动态化集成研究已成为趋势,为生态保护、农业升级和应对全球气候变化等提供科学基础。
能量流动与生态系统功能
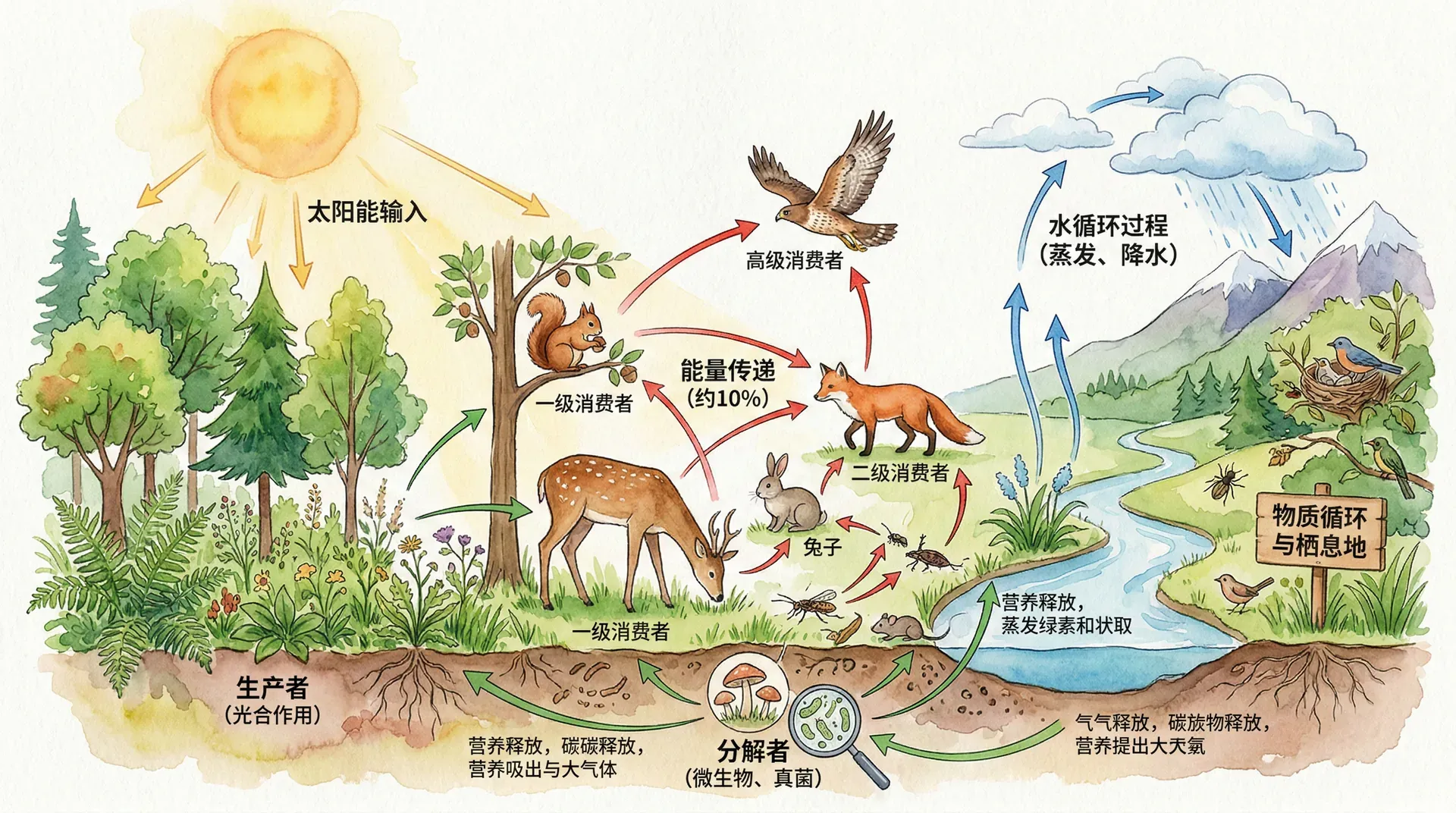
能量流动不仅是生态学的基础理论,还直接影响着生态系统的多种功能。理解能量流动规律,有助于更科学地管理和保护生态系统。比如,在畜牧业生产中,约10%的能量可以从低一级传递到高一级,即生产1千克牛肉需消耗约10千克饲料。因此,从能量利用角度看,人类直接食用植物比转化为动物性食品效率更高。但由于人类无法直接利用多数草地植物,草食动物的存在让这些能量得以“转化”为我们可用的蛋白来源。
在中国西部牧区,合理放牧依赖于草地的净初级生产力(NPP)。假如每公顷NPP为4000千克,按10%的能量传递效率,理论上支持400千克家畜生物量。但出于生态系统稳定和可持续考虑,实际放牧量应该控制在理论值的50-70%,以防止草地退化。
过度放牧会打破草地能量平衡,导致植被衰退、土壤侵蚀及荒漠化。因此,必须根据NPP和能量流动效率科学确定家畜数量。
能量流动分析同样适用于城市。北京等大城市生态系统,能量主要来自化石燃料和电力,而非自然太阳能输入。城市生态系统通过增加绿地,提升植被覆盖率,不仅有助于太阳能利用,还可缓解热岛效应,改善人居环境。
在生态恢复工程中,如黄土高原退耕还林还草,提高植被覆盖直接带动NPP的提升。例如,退耕区NPP由每平方米5000千焦耳增至18000千焦耳,增强了水土保持和生物多样性。
此外,能量流动研究还揭示了生态系统稳定性的机制——复杂的食物网意味着能量流动路径更多样,系统抗干扰能力更强。当某条通路受阻时,能量可通过其他通路维持系统功能。因此,保护物种多样性实则是在维护生态系统能量流动网络的完整性。
本节练习
1:某草原生态系统中,牧草年净初级生产力为每公顷6000千克(干重),能量含量约为18000千焦耳/千克。若能量传递效率为10%,请计算该草原理论上可以支持多少千克羊的生物量?假设羊的能量含量为20000千焦耳/千克。
答案:
计算牧草的总能量:
2:在某森林生态系统的食物链中:橡树 → 毛虫 → 山雀 → 鹰。已知橡树固定的太阳能为2×10⁷千焦耳/公顷/年,如果每个营养级的能量传递效率分别为10%、12%、15%,请计算鹰每年可以获得多少能量?
答案:
毛虫从橡树获得的能量:
3:下表是某湖泊生态系统四个营养级的能量流动数据(单位:千焦耳/平方米/年):
请完成表格中的空缺数据,并计算浮游植物到浮游动物的能量传递效率。
答案:
根据能量平衡原理:同化量 = 呼吸消耗 + 流向下一营养级 + 流向分解者
计算浮游植物流向下一营养级的能量
流向浮游动物的能量为 千焦耳。
4:某农田生态系统中,农作物的总初级生产力(GPP)为50000千焦耳/平方米/年,呼吸消耗占GPP的40%。在收获季节,农民收获了占NPP 60%的农作物,其余40%作为秸秆还田。请计算:(1)农作物的NPP;(2)农民收获的能量;(3)秸秆还田的能量。
答案:
- 计算NPP
5:为什么大多数食物链的营养级不超过4-5级?请从能量流动的角度解释这一现象,并说明这对生态系统结构有什么影响。
答案:
食物链长度受限的主要原因是能量传递效率低。
(1)能量递减原理:
能量在营养级间传递时,效率通常只有10%左右,即每传递一个营养级,就会损失约90%的能量。经过4-5个营养级后,剩余能量已经非常少。例如,如果生产者固定10000单位能量,按10%传递效率计算:
-
第一营养级(生产者):10000单位
-
第二营养级:1000单位
-
第三营养级:100单位
-
第四营养级:10单位
-
第五营养级:1单位
到第五营养级时,可用能量已不足以维持生物种群的基本生存需求。
(2)对生态系统结构的影响:
① 限制了顶级消费者的数量和体型:由于可获得的能量有限,顶级捕食者的种群数量通常较少,个体较大以提高捕猎效率。
② 影响了生态金字塔的形态:能量金字塔始终呈正金字塔形,底部宽大,顶部狭窄。
③ 决定了生态系统的承载力:可利用能量的多少直接决定了生态系统能够支持的生物量和物种数量。
④ 塑造了生物多样性格局:低营养级的物种多样性通常高于高营养级,因为有更多能量可供利用。
知识点:本题考察对能量传递效率及其生态学意义的综合理解,要求学生能够将理论知识应用于解释生态现象。
6:在中国东北地区,近年来野生东北虎数量有所增加。从能量流动的角度分析,保护东北虎这种顶级捕食者需要保护多大面积的森林生态系统?请说明你的分析思路。
答案要点:
(1)分析思路:
东北虎作为顶级捕食者,位于食物链的最高营养级。根据能量传递效率约10%的规律,支持东北虎生存需要庞大的生态系统基础。
(2)能量需求估算:
一只成年东北虎每天需要摄入约5-7千克肉类,年消耗约2000千克。假设能量含量为千焦耳/千克,则年能量需求约为千焦耳。这些能量来自野猪、鹿等猎物(第三营养级)。
按能量传递效率逆向推算:
-
支持东北虎需要的第三营养级能量:千焦耳
7:比较陆地生态系统和水生生态系统在能量流动方面的主要差异,并解释为什么海洋渔业的生产力通常比陆地畜牧业更依赖于较短的食物链。
答案要点:
(1)主要差异:
① 生产者特征不同:
陆地生态系统的生产者主要是大型植物(如树木、草本),个体大、寿命长、生物量积累多;水生生态系统的生产者主要是浮游植物,个体小、寿命短、周转快。
② 生物量金字塔形态:
陆地生态系统通常呈正金字塔形;水生生态系统可能出现倒金字塔形,即某一时刻浮游植物的生物量可能小于浮游动物,但全年累计的能量产出仍大于消费者。
③ 能量周转速度:
水生生态系统的能量周转速度快,浮游植物的世代时间通常只有几天到几周;陆地植物的世代时间从几个月到数十年不等。
(2)海洋渔业与食物链长度:
海洋渔业更依赖短食物链的原因:
① 能量损失:每增加一个营养级,损失约90%的能量。海洋中许多经济鱼类(如金枪鱼、鲨鱼)位于较高营养级,如果食物链过长,可利用的能量极少,无法支撑大规模渔业生产。
② 生产效率:人类直接捕捞低营养级鱼类(如鲱鱼、沙丁鱼)或养殖以浮游生物为食的鱼类(如鲢鱼、鳙鱼),能够获得更高的能量利用效率。中国的淡水养殖业主要养殖鲢鱼、鳙鱼等滤食性鱼类,它们直接以浮游生物为食,食物链短,能量利用效率高,因此产量大。
③ 对比陆地:陆地畜牧业中,草食动物(如牛、羊)只经过一次能量转换,而海洋中许多经济鱼类需要经过2-3次甚至更多次能量转换,导致生产效率降低。
(3)实践意义:
这一认识指导渔业生产向“短食物链、高效率”方向发展,如发展低营养级鱼类养殖、推广滤食性鱼类、控制过度捕捞高营养级鱼类等。
知识点:本题考察对不同生态系统能量流动特点的比较分析能力,以及将理论应用于解决实际生产问题的能力。