生命的本质
当我们抬头仰望星空,望着无边的宇宙时,难免会生出疑问:“我们是谁?生命究竟意味着什么?我们存在的意义在哪里?”这些疑惑自古以来便萦绕在人类的心头,无数哲学家、诗人和科学家都曾试图给出自己的答案。在很长一段历史时期,人们对生命的解释建立在神话、传说以及丰富的想象之上,或赋予超自然的力量,或寄托在模糊的信仰之中。
然而,到了近代,随着科学的进步,一位伟大的思想者的出现彻底改写了我们对生命的理解。他用严密的观察、扎实的证据和清晰的逻辑,为这个古老问题提供了全新的视角。正是他的理论,成为理解生命奥秘的关键,使得人类第一次能够用科学的方式去揭示生命的本质。这场观念的变革,不仅让我们走出盲目的迷雾,也让我们得以用理性的眼光审视自己的起源和未来。
进化论
1859年以前,人类对于“我们从哪里来、为什么存在、生命为何如此多样”这些问题,大多只是基于神话传说、宗教解释或单纯的猜测,没有科学的证据和体系。每个古老文明都有自己的生命起源故事——有的讲述女娲造人,有的相信神灵赋予生命,甚至有人认为人类与自然万物毫无关联。可是,这些解释都无法系统地解答生活中对生命现象的好奇与疑惑。
这一切直到十九世纪中叶终于发生了革命性的变化。英国科学家查尔斯·达尔文在1859年发表划时代的著作《物种起源》,首次提出了“进化论”这一大胆假设。他认为,所有现存的生物,包括人类,都是在极其漫长的时间里,通过自然选择与遗传变异逐步演化而来。生物体并非一成不变,而是在不断适应环境变化的过程中“筛选”出了最适应现状的特性,并且将这些特性传递给后代。这种不断变化和适应的过程,就是生命多样性的根源。
进化论的重要性,不亚于哥白尼和伽利略发现地球围绕太阳运转。正如“日心说”打开了天文学和物理学新纪元,达尔文进化论不仅揭示了生物多样性的成因,也成为我们理解一切生命现象以及自我定位的科学基石。它解释了生命为何会有如此丰富的适应形态,还让我们第一次理解了生命背后隐藏的自然规律。
基因的自私本性
现在让我们从一个全新的角度来思考生命的本质。假如我们要了解一个人在深圳的科技公司中如何生存下来、并且取得事业上的成功,我们往往会去分析他在这个高度竞争环境中的表现。通过观察他的行为与结果,我们会推测他一定具备某些独特的优势:比如卓越的专业技能、良好的人际沟通能力、或者足够强大的抗压和适应变化的能力。这些特质帮助他在激烈的竞争中脱颖而出,获得更多机会、晋升甚至领导他人。
但你有没有想过,这样的道理同样也适用于基因的世界?在进化论的视角下,生物体其实只是基因的“载体”或“工具”。我们每个人,以及世上所有的生物,其实本质上都是由基因“制造”出来的复杂机器。基因需要依托一个载体(也就是生物个体)来完成自身的复制与传递。能够长时间存活、并且代代相传的基因,必然具备某种能够提升自己“复制概率”的关键特征。这个特征就是我们所说的“自私性”——基因会倾向于采取任何有利于自身留存与传播的行为策略。
这里说的“自私”其实并不是通常意义上的道德评判,比如自利或自我中心。恰恰相反,这只是对基因运作模式的科学描述。所谓“基因的自私性”,指的是基因本能地倾向于最大化自身在未来世代中的出现频率——也就是说,采取任何能提升自己被复制和遗传到下一代的策略,哪怕表面上看起来这个行为“无私”。
事实上,基因的自私本性,常常在生物个体的行为层面表现为自我保护、竞争、资源独占等自私行为。但奇妙的是,基因有时也会通过促进“利他行为”来实现自己最终的“自私”目的。例如,在某些特定的状况下,一只动物对同类表现出帮助、牺牲甚至自我牺牲的行为,实际上可能是因为这种行为有助于让带有相同基因的个体获得更高的生存或繁殖机会。换句话说,基因为了“自保”,有时也会间接推动“合作”或“利他”的出现!这种看似矛盾的机制,其实揭示了自然界行为的复杂性与进化的多样路径。
我们或许可以用最直观的数据来说明基因自私性对种群的影响。假设一个种群内存在“合作基因”和“自私基因”两种遗传策略。合作基因鼓励个体付出代价帮助同伴,而自私基因则让个体优先考虑自身利益。如果没有特殊形式的机制鼓励合作(比如亲缘选择、互惠利他或强制性惩罚),哪些基因最终会占据上风呢?
这个图表直观地揭示了一个残酷但真实的进化法则:如果没有有效的机制来保护和奖励合作,那么在漫长的演化过程中,“自私基因”往往会因为更高的个体生存和繁殖效率而逐渐取代“合作基因”,占据整个种群的主导地位。这也解释了为什么在自然界中,纯粹的、无条件的利他主义极为罕见,而大多数合作都伴随着复杂的制衡和反馈机制。理解这一点,对我们认识生物本性、社会行为的演化,有着至关重要的意义。
科学描述与道德判断的区别
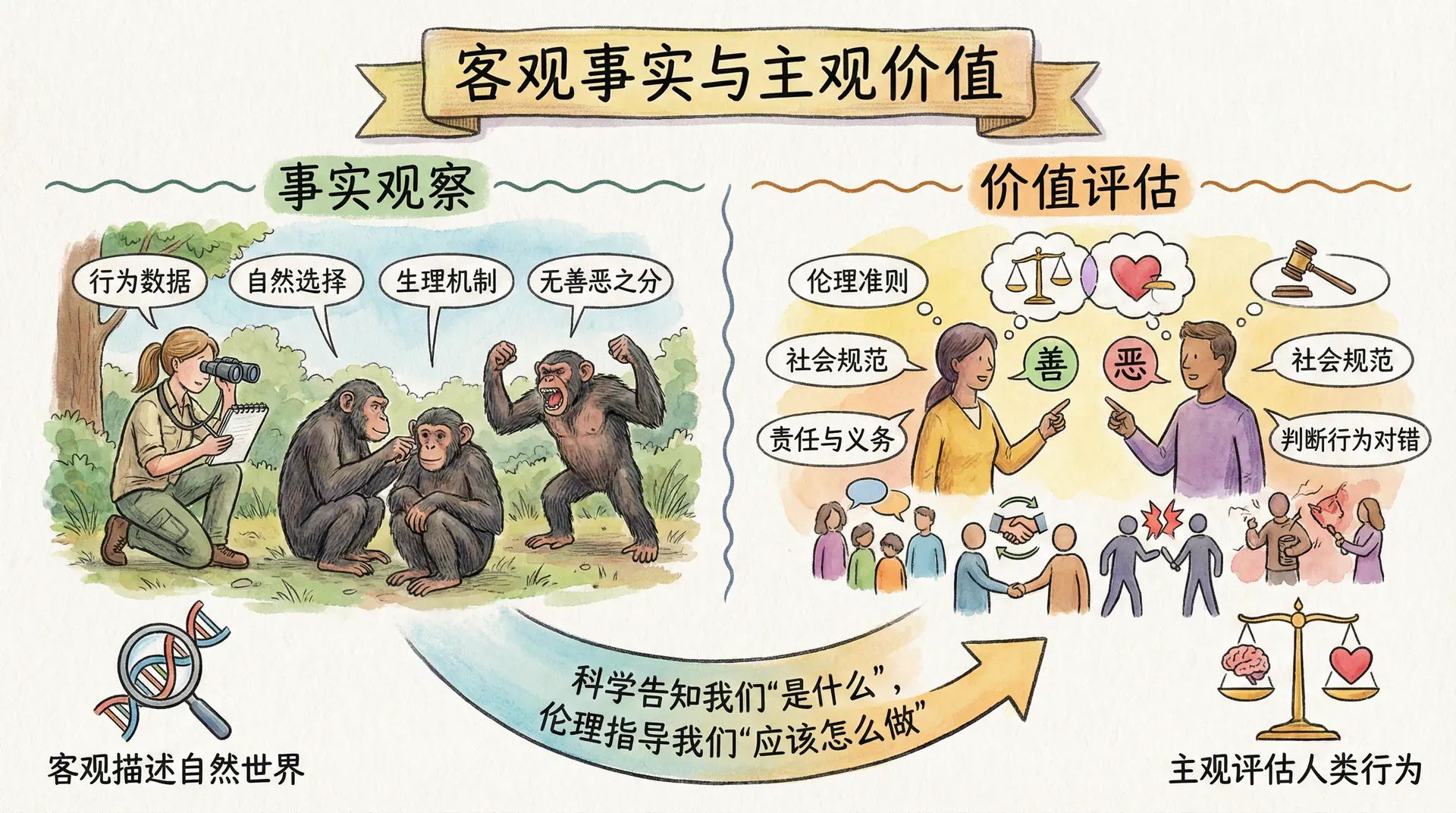
在我们继续深入探讨进化论所揭示的人性与行为时,必须首先区分两种截然不同的讨论方式——一种是对自然界本质的科学描述,另一种则是社会中关于价值观和行为准则的道德判断。
科学描述关注的是事实,是对现象的客观解释。例如,当我们说“基因具有自私性”,这只是指出生物演化中某些规律,并非鼓励人类社会仿效基因的自私逻辑。正如物理学揭示了“重力作用”,但我们不会因此得出“人应该彼此下拉使其坠落”的道德结论。理解这一点十分关键,因为如果混淆了科学描述和道德判断,容易铸成“自然主义谬误”——误以为“自然如此”即“应当如此”。
事实上,正是因为我们理解了基因的自私机制,才更能察觉建立合作和利他行为在社会中的重要性。换句话说,科学为我们揭示了本性,而道德和文化则赋予我们改变自我的可能性。人类与其他生物最大的不同之一,就是我们可以通过教育、伦理、制度设计等“后天力量”,有意识地调整甚至对抗某些天生倾向。
我们作为人类,拥有独特的思考和选择能力,这让我们能够跳脱基因的“指令”,用文化和理性不断塑造自身。正因如此,只有人类才能主动塑造未来、建立超越利己本能的文明社会。
理解进化机制不是在为“自私”辩护,而是让我们看清需要在哪些地方付出额外努力。我们欣赏英雄、坂雪救人的故事、赞美无私奉献,恰恰因为这些行为超越了本能,不是轻易就能实现的。
先天与后天的辩论
关于人类行为,长久以来都有一个激烈纷争的问题:我们的性格、智力、情感等,是更多由基因决定,还是由成长环境和文化影响塑造?这就是著名的“先天(nature)与后天(nurture)”之争。
一些学者和公众倾向于将一切归咎于先天遗传,认为我们的行为和能力基本是“写在基因里”的;而另一些人则强调后天环境和文化教育的决定性作用,甚至主张“人之初,性本善恶全赖教养”。但真实世界远比这两极化看法复杂得多。
在中国的独生子女政策中,几十年间,一代独生子女成长起来。有人发现在行为模式、处事方式、家庭责任感等方面,独生子女与多子女家庭的孩子存在一些显著差异。这种差别是因为基因突变了吗?明显不是。这里的“行为模板”是在特定社会政策和家庭结构中塑造出来的,环境起了决定性作用。
但如果我们把目光放到更多样、极端或跨文化的情形下,会发现每个人的成长轨迹,既受限于生物学赋予的潜能(如气质、智力、心理倾向等),又受环境和文化的持续塑造。基因为我们预设了行为和发展的基本“地基”,而文化则像盖楼一样不断加码与精雕细琢。
值得一提的是,即使某些不良倾向(如攻击性、自私等)在进化过程中有“遗传基础”,这也绝不代表我们只能任其自由发展。实际上,人类文明和教育的目标之一,就是用制度和道德力量校正、引导甚至改写进化带来的“本能冲动”。我们之所以能有条件建立互助、宽容、社会责任、道德感,这背后既有文化累积,也有理性自由选择的力量。
行为的科学定义
说到生物行为,我们需要抛开日常的价值判断,采纳标准化、便于分析的科学定义。这里有两个最基本也是最常引发争议的概念:利他行为与自私行为。
在生物学领域,利他行为是指:某个个体的行为会提升其他个体的生存或繁殖概率,却降低了自己同样的概率。与之相对,自私行为是指:某个个体的行为牺牲他人利益以换取自身生存或繁殖概率的提升。
请注意,科学定义中的“利他”与“自私”只涉及行为结果,而不涉及动机或主观意图。比如,一只鸟喂养幼鸟,这一行为归为“利他”,无论它是出于本能还是出于“爱”。科学关注的是因果关系和客观效应——谁从中获益、谁付出代价。
这样的定义看似冷酷简单,但恰恰是它让我们在宏观层面精准量化、比较不同策略的长期影响。在自然界,甚至百分之一、千分之一概率差异,经过数十代的积累都会让“赢家”与“输家”拉开巨大差距。
假设一个种群中有两种行为策略,一种让个体的生存率每一代提升1%,另一种则导致生存率每一代降低1%。一代两代也许变化微弱,但五十代之后呢?我们用真实数据展示这一影响——
从这个对比图可以看出,即使只是1%的差异,通过长期叠加都会产生“雪球效应”——五十代之后,优势策略带来的种群数量可能是劣势策略的近3倍以上。这正是进化过程能够如此高效塑造各种特征的核心动力来源。
这些科学事实不仅让我们敬畏自然的复杂与力量,也提醒我们:微小改变有可能决定整个种群乃至社会的命运。从中我们学会谦逊,更激发了我们对于人性、文化、社会塑造的创造性思考。
自私行为的实例
下面我们将详细探讨自然界中自私行为的多样化表现,以及它们背后的生物学意义。不仅仅局限于书本或国外的案例,而是紧密结合我们身边可见的生态环境。
鸟类的竞争
在我国众多湿地保护区,苍鹭的行为为我们提供了研究自私行为的绝佳窗口。研究者发现,苍鹭喜欢在水边相对密集地筑巢。在雏鸟孵化后,成年苍鹭不仅会悉心照顾自己的后代,还会密切关注邻近巢穴的动静。如果邻巢的成鸟外出觅食,有的苍鹭会趁机靠近,并袭击别家幼鸟。这一行为带来的直接好处是:它可以较为容易地获得高营养的食物,而无需冒险飞到遥远的水域觅食;间接的影响则是弱化了未来的竞争——减少了非血缘后代的存活几率。这种行为从表面看很残酷,但实际上却有助于提升自身基因的传播概率。
事实上,这种现象在其他鸟类中同样存在。例如在中国北方,一些燕雀类小型鸟类也会发生“偷蛋”或夺巢的行为。禽类通过夺取他巢资源,能显著提高自身及其后代的存活率。这些策略都围绕着“自我最大利益优先”展开,是典型的自私行为。
社会性昆虫的现象
转向常见的社会性昆虫,蜜蜂的自私行为同样不可忽视。有人可能认为,工蜂蜇人时即使死亡也在保护蜂巢,是一种利他行为。事实上,如果我们把观察尺度拉大,从基因层面审视,这恰恰是自私性的高级表现。
让我们具体分析:
如上表所示,如果工蜂仅考虑自身安全而逃避危险,则整个蜂群面临更高风险,影响了拥有相同基因的姐妹的生存。如果蜂群没有防御机制,种群难以延续。主动进攻的“牺牲”表面利他,本质恰是“家族基因自私性”的体现。这种行为经过数千代进化,成为社会性昆虫维系群体生存的关键机制之一。
此外,在田间地头观察蚁群,有时也可以发现类似的激进防御行为。当蚂蚁巢穴受到威胁,前方的“战士蚁”会冲在最前线,牺牲自己守护巢群。细查下来,这些蚂蚁大多与蚁后有密切血缘关系。换言之,个体的“牺牲”只要能促进近亲基因的存留,便是自私性的又一种变体。
生存竞争与资源独占
在我国南方的一些农田区域,灰喜鹊等鸟类会为了争夺有限的筑巢区或食物资源,采用极端排挤甚至暴力驱逐的方式,赶走其它竞争者。这是典型的通过减少他人机会来提升自身生存概率和后代表现的自私行为。冬季时,鸦类也会埋藏食物,有时候甚至会监视同伴并在对方离开后悄悄“盗挖”,完全不考虑他人的利益。
企业竞争的类比
如果我们把视角从动物界转向人类社会,其实也有很多可以对应的现象。例如在深圳的科技创新园区,创业公司在获得融资的竞争中经常会做出极具针对性的“防守”动作。假如某家公司在与投资人洽谈时,另一家公司突然低价推出相似产品,或在行业会议上高调发布新计划,这些行为本质上是为了提升自己赢得投资青睐的几率。虽然短暂来看有利于个体企业,但长期看或许会损害行业整体利益、甚至引发恶性竞争,这与生物界“自肥”而无意于集体利益的策略如出一辙。
在自然界,无论动物还是昆虫,自私行为并不等于“道德缺陷”。它是进化压力下个体为优化自身基因传播效率所自发采取的客观策略。这些策略有时候会导致竞争、冲突、短期损失甚至牺牲他者利益,但它们共同支撑起了生物多样性及其复杂生态系统的运转。
我们需要理性理解自私行为,并通过科学观察与分析,明白其背后的生物学逻辑,这样才能更好地借鉴自然界的规律,用于指导人类社会管理和制度设计。
利他行为的表象
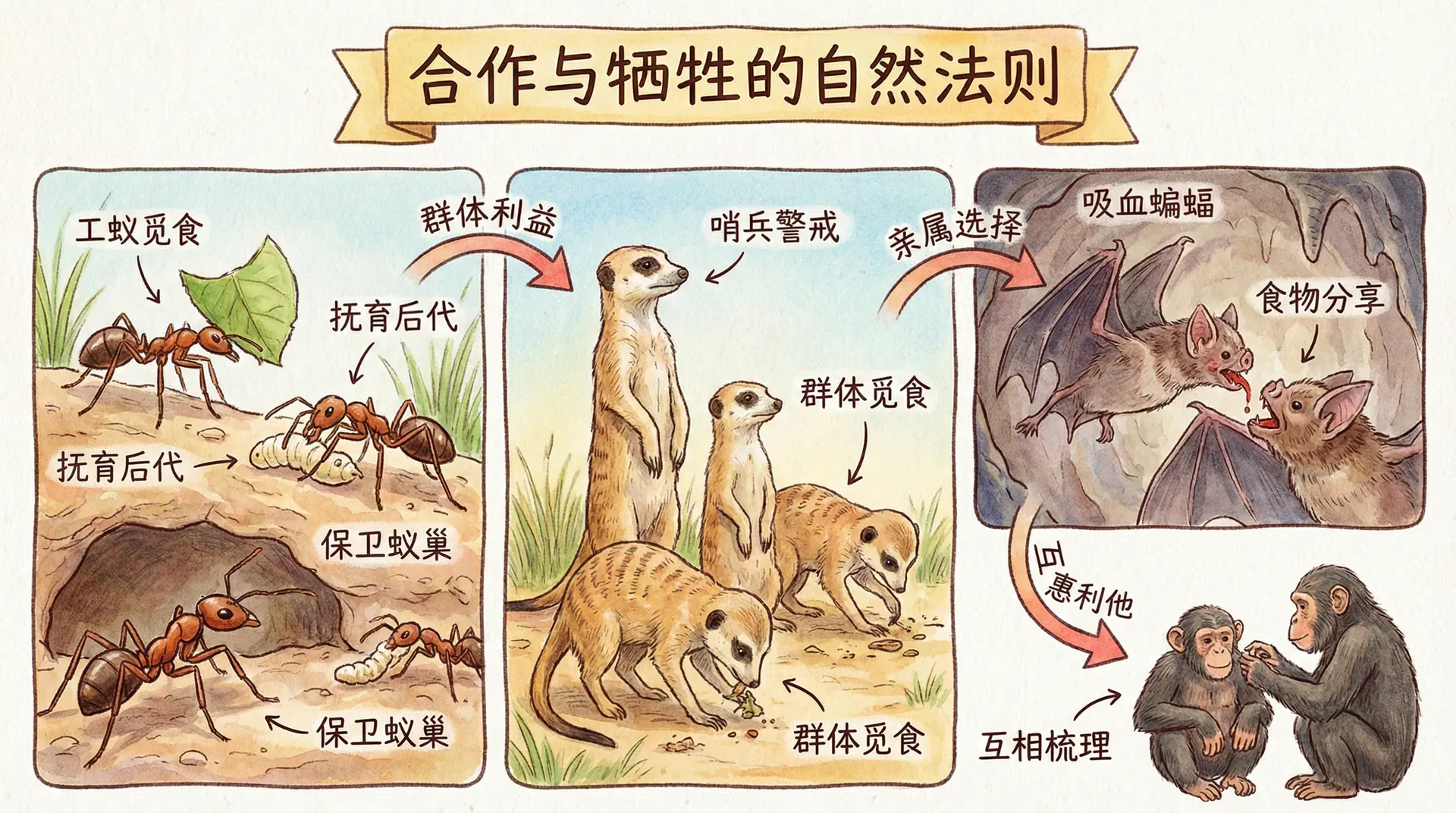
我们已经了解了“自私行为”在自然选择中的作用,那么现在让我们转向另一面:看上去是利他的行为。利他行为为生物进化史增添了不少复杂性和争议,看似“牺牲自己、成全他人”,但生物学家们发现,背后的动力常充满了进化的智慧。
父母的投资
在自然界中,最普遍、最直观的“利他”行为来自父母对子女的照顾。几乎所有的鸟类、哺乳动物,都会极为卖力地照顾自己的后代。比如鸟类每次觅食都要飞行数公里,为雏鸟衔回食物;哺乳动物在遭遇天敌时宁可自己受伤,也要保护幼崽安全。这些行为在野外观察非常常见,无论是在森林、公园,还是农田周围都能见到感人的场景。
以长江流域白鹭为例,研究人员观察到在巢区内,当蛇或其他捕食者逼近时,成年白鹭常常采取“佯装受伤”策略:它假装翅膀折断一瘸一拐地在地上跳跃,吸引捕食者的注意力,表现得像一个“容易捕获的猎物”。捕食者通常会被诱惑,转而追逐成年鸟,放弃了对鸟巢的威胁。等到危险消除,成年白鹭则会突然振翅高飞,安全逃脱。这一系列动作不少动物学家都亲眼见过,称为“转移注意力”行为。
这种行为表面上看是父母为保护幼崽不惜以身犯险,似乎无私奉献,但从进化生物学、尤其是基因角度分析,这是极为高明的策略。成年鸟冒险其实是为了保护拥有自身基因的后代,从而最大化基因的传递概率。换句话说,表面利他,本质仍然是“基因自私性”的体现—类似行为在人类和其他哺乳动物中也很常见(如母猴舍身对抗猛兽、哺乳期家犬猛烈地顶撞入侵者等)。
不仅如此,不同物种还有更加“极端”的利他型育幼策略。例如在部分鲤科鱼类中,父母会将几乎全部体能消耗在产卵和护卵上,甚至在孵化后体力耗尽、失去存活机会。这样的“牺牲”行为与基因的延续和“亲代投资理论”密切相关。
预警行为
除了对子女的照顾之外,群居动物之间也经常有看似“舍己为人”的行为。比如在云南的部分森林、农田周边,经常能观察到"麻雀预警"的现象。在鸟群觅食或休息时,总有几只麻雀格外警觉。当发现远方有老鹰或其它猛禽接近,它们会用特殊的叫声(如刺耳的“啾啾”声波)提醒同伴。整群鸟在收到警告信号后会瞬间四散飞开,大幅提高了整个群体的生存概率。
发出警报的鸟表面上承担了额外的风险:叫声不仅暴露了自己的位置,还可能吸引捕食者先攻击它自己。这种举动乍一看无疑是利他的,但如果深入研究,被警报所庇护的是谁?答案是——鸟群中往往包含大量发出警报个体的亲属。尤其在同一窝孵化或同时间成长的个体之间,这种亲缘关系更为密切。
进化生物学的“亲缘选择理论”(kin selection)指出:当利他行为的受益者与施予者有较高的基因共享度时,行为所付出的代价完全可以被基因传递概率的提升所弥补。这就解释了为何类似“报警牺牲”的举动会在自然界广泛存在。
为便于理解,让我们继续用一组数据来分析这个过程:
我们可以看到,发出警报个体自身的生存率会有一定提升,但鸟群整体的生存概率因警报而获得了“大幅度”上升。假设鸟群中20%是报警鸟的近亲,那么即使个人冒险,整体基因的传递几率依然更高。从基因的长远利益来看,报警行为也是一种“理性自利”,它通过帮助亲缘个体间接实现了“自我基因”的扩展。
实际上,类似的“利他”例子在中国常见的田鼠、旱獭等啮齿类动物中也广泛存在。例如新疆草原旱獭哨兵会站在高地观察,一旦有天敌靠近即刻发声预警;高原鼠兔同样能够通过“打哨—躲穴”协作,大大降低被捕食的风险。这些都说明,“利他行为”的本质,其实与进化过程中的理性自我利益息息相关。
群体选择的误区
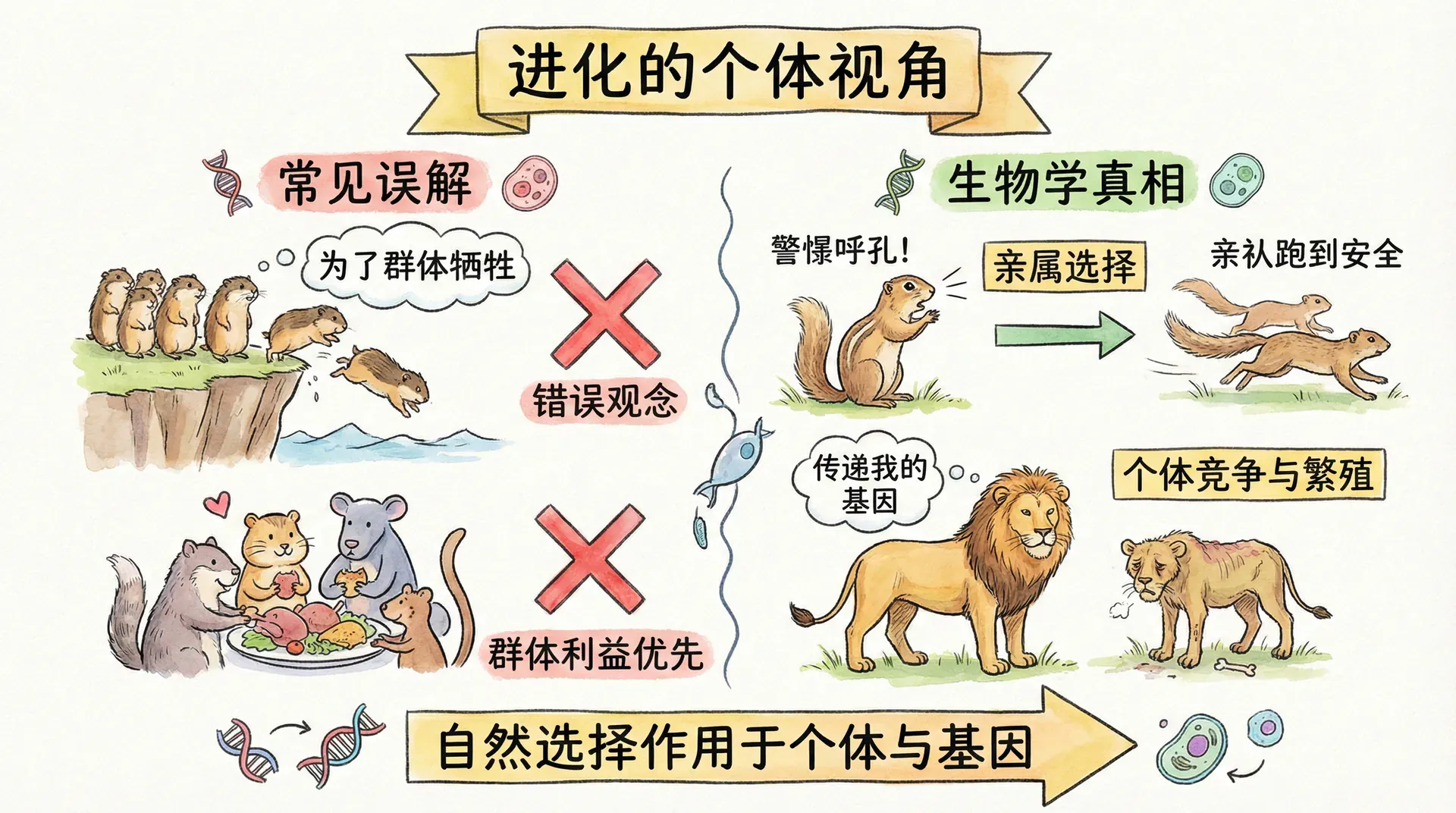
在接下来的讨论中,我们要深入剖析一个在进化生物学中长期存有争议、且常常被大众误解的重要议题:究竟动物的行为会不会出于“物种整体的利益”而行动?为什么看似为群体或物种“牺牲自我”的行为,其实并非表面看起来那样简单?
事实上,许多人在人类文化和道德观念的影响下,容易将动物行为类比为“为集体、为物种”的高尚选择。这种观点看似合乎伦理,道德上也令人向往,然而从现代进化理论的角度来看,它并不符合自然选择运行的根本逻辑。
逻辑问题与思想实验
为了更清楚地理解,我们不妨以一个经典思想实验为例,来剖析群体选择理论的内在矛盾。假设存在两种完全不同的种群策略:
种群A:其成员都高度利他,愿意为了集体利益牺牲自身,即在资源短缺时主动减少繁殖,甚至自愿饿肚子,只为让整个种群能持续下去。
种群B:所有个体都以自身利益最大化为目标,遇到资源紧缺时仍各自竞争、尽量繁殖和摄取食物,不考虑集体的存续。
乍一看,种群A似乎更具“理智”与长远眼光,因为它们能防止集体的资源枯竭,似乎为整个种群争取到了更好的未来。但让我们更进一步思考实际演化会发生什么。
假如在种群A中某一代出现了一个“叛徒”——即突然出现一个只顾自己、不愿牺牲的个体。当所有成员为集体利益自我约束时,这个叛徒就获得了更多的生存与繁殖资源,能够养育出更多后代。而且,这种“自私”性状会被它的后代复制,于是“自私”个体越来越多,逐步取代了那些老实利他的成员。
通过下图感受到这个变化过程:
纵使在起初,群体中只有极少数叛徒,随着一代代繁衍,这些“自私”个体总会迅速占据遗传优势,从而稀释甚至淘汰“利他”基因。由此可见,群体选择理论在根本上难以抵抗个体层面的自私行为——这是内在逻辑决定的。
中国农业改革
上述进化现象不仅存在于自然界,在人间社会历史上也屡见不鲜。比如中国上世纪80年代著名的农村“人民公社”到“家庭联产承包责任制”的转变,就是一个高度真实的例子。
在人民公社集体生产体制下,理论上每个人都应承担集体责任,为大家的共同利益工作。但实践中由于个人努力与个人收益之间缺乏直接关联,导致“多劳未必多得”,人们普遍工作积极性降低。有人偷懒,损失由集体承担;有人勤奋,收益也被集体平摊。因此,在这样的激励机制下,“偷懒不干活”反而成了理性人的选择。
而改为“家庭联产承包责任制”以后,个人的辛勤和成果直接挂钩。激励到位,每个人都倾向于自利地付出更多劳动,农业生产力便随之大幅度提升。这恰恰揭示出,如果没有足够机制协调、绑定个体与集体利益,单靠“集体意识”很难持续。现实与进化一样,对自身利益的优先考虑往往是最有力的动力。
关于“物种选择”的进一步荒谬推演
我们再来分析群体选择理论的另一层致命弱点:时间尺度与选择压力。
理论上,群体选择理论的支持者认为,充满利他行为的群体能够存活下来,而充满自私者的群体会被淘汰。但是,进化选择在不同层次上,速度和强度是天壤之别的:
我们可以看到,个体内、个体间的竞争作用几乎“即时”发生并迅速见效,选择压力极大。而群体间的淘汰和物种间的竞争,则需要很长的时间尺度,且作用弱得多。换句话说,即使某个群体长期来说因太“自私”最终被淘汰,但还没等这一结局到来,这个群体早就被自私个体从内部侵蚀了。
有些人甚至将“选择”层级无限上推:如果物种也是竞争单位,那么是不是所有哺乳动物都应该联手,不再自相残杀,对抗爬行动物或者鸟类,因为它们属于更大的“纲”?再或者,各门动物为了对抗外界威胁,跨界合作?而事实上我们从未见过狮子因“哺乳动物同盟”而放弃猎杀羚羊,老鹰也不会因为同属“鸟类”而宽恕麻雀。
这正好反映了群体选择理论的荒谬与局限性:随着选择层次上移,选择压力越来越弱,约束力越来越小,远远无法对抗个体、基因层面的强大动力。演化的真正“战场”,仍然是在个体甚至是基因的层级上,而非玄而又玄的“物种利益”。
群体选择的误区提醒我们,要警惕那些看似“高尚”、实则经不起分解分析的观点。生物不会因为“物种或生态系统”的整体利益而自行约束行为,进化机制依然是“具体且微观”的自利与选择。
基因层面的选择
那么,选择究竟在哪个层次上发生?如果说自然界中的生存斗争有一条根本线索,这条线索最终会指向基因本身。实际上,自然选择最基本、最直接的单位,是基因。
这个观点听起来或许有些激进,但它恰好是最符合逻辑和事实观测的方式。并不是说个体、种群或者群体就不重要,而是在解释进化、遗传和生命多样性最根本的原因时,把视角聚焦到基因层次,才能让很多现象变得清晰、合理。
为什么我们要从基因看世界?
例如,基因好似一条条“信息指令”,这些指令能精确地传递下去。当一个人去世,他的基因很可能已经通过生儿育女等方式,部分或者整体传给了下一代。如果这个人从未留下后代——他的基因“走到了终点”;但那些顺利被复制且留存于后代中的基因,就有机会继续“旅行”下去。也即,基因不像个体那样会终结,而是在生物世代之间长久流传。
为了更清楚地理解,我们可以用一幅建筑图纸来打比方:每一个生物个体就像是一栋大楼,而基因就是设计方案和施工蓝本。大楼(个体)可能会毁坏或倒塌,但一旦图纸(基因)广为流传、不断用于新楼的建造,这套图纸就能在城市里延续不朽。同理,一个好的基因通过不断复制自己,能够在自然世界中持续存在,而不适应环境的基因则逐步消失。这就是自然选择在基因层次的本质。
基因可以实现长久的延续并精准复制,而个体生命有限、只能通过繁衍传递遗传信息。两者虽相互关联,但在自然选择中侧重点明显不同——基因层面更注重长远影响和稳定传递,个体层面则更代表立即的生存表现。
基因的运作揭示生命奥秘
在基因层次上思考生命,可以帮助我们很好地解释许多看似“反常”的现象。例如:
在中国北方农村,蜜蜂养殖早已成为重要产业。大家知道,工蜂在蜂巢受到威胁时,会毫不犹豫地牺牲自己。为什么?因为牺牲工蜂自身,可以保护蜂巢(包含许多与自己有极高基因相似度的“姐妹”);这样一来,自己的基因间接得到了最大程度的保存和繁衍。
再比如,为什么许多动物父母对自己的子女关怀备至,甚至不惜犯险呢?从基因角度,这一行为就是典型的“基因自我复制动力”:父母与子女拥有一半完全相同的基因,照顾后代就是优先保证自己的基因能顺利传递下去。
然而,基因层面的选择并不意味着每次都会做出“善良”或“合群”的决定。在一些鸟类中,食物短缺时会发生“雏鸟互吃”甚至父母主动淘汰最弱小雏鸟的现象。从人类情感看这是残酷的,但基因“考量”的是整个基因组后代的成功率——把资源集中给最有可能存活的雏鸟,是确保基因最大化留存的策略。
从基因视角审视身边的社会关系
在我们的日常生活里,基因层面选择的痕迹也处处可见。比如为什么人们在亲属间愿意互帮互助,而在陌生人之间更多表现出防备和保留?这是基因相同或相近,利益趋同的自然结果。同时,这种倾向也解释了“中国式家庭”里父母对子女无条件付出的文化特征——从基因角度,这些“无条件的爱”其实是优选了遗传自身特征的自然机制。
当然,人类发展至今,已超越了很多基因本能。我们建立社会规范、道德约束,甚至创立教育体系,激励人们为更广阔的群体和无血缘关系的人合作。这也说明,人类具备打破基因“脚本”的意识和能力。
总结
为什么我们要对这些深刻的自然规律进行反复推敲?因为它们不仅能帮助我们洞察生命奥秘,更帮助我们理解自我和他人的一举一动。
试想中国历史上的“互助组”,一开始是小范围几家人互帮互助,后来发展到生产队、村集体。当集体利益与自身利益一致时,合作顺畅;但如果两者出现冲突,个体往往倾向于优先考虑自己或者亲族成员。这不仅仅是社会环境的产物,更有深刻的生物进化基础。
很多时候,我们发现合作的实现相当困难,尤其是当涉及陌生人或者与我们没有直接联系的人的时候。正因如此,社会才需要建立制度、文化和法律,对个体的“天然自利”加以调节,从而实现更广泛的合作和共赢。
此外,我们还需意识到,人类是地球上唯一可以清醒意识到自身基因局限,又能主动为整体、为理想、为更高层次目标而影响甚至改变本能倾向的生命。我们会主动帮助弱者,会为道义放弃眼前利益。在生态保护、支援灾区、志愿服务等各类场景,中国社会到处涌现“超越基因脚本”的壮丽行动。
人类是唯一能够认识到自己基因程序,并有能力进行某种程度上“反抗”或超越的生命形式。这并不是弱点,恰恰是我们最宝贵的特质。
我们对基因层面选择的理解,不是说“自私就是天经地义”,更不是为一切短视行为找借口。相反,这帮助我们警醒:美好的社会需要有意识地设计和维护,需要勇气和智慧去“逆流而上”,创造出超越生物本能的合作与责任感。这种付出和努力不会因为困难而失去价值,恰恰因为稀少和可贵而更值得敬重。
未来我们会继续深入探索生命诞生的奇迹、基因运转的奥妙,以及复杂生物形态如何由简单到复杂逐步发展。不论我们挖掘哪些知识,都要记住:理解自然选择最重要的出发点和归宿,就是聚焦基因层面。这正是揭开自然之谜的“钥匙”,也是生物学学习的恒定指南。