生命之树的故事
站在云南西双版纳的热带雨林中,你能听到长臂猿的鸣叫;攀上青藏高原,藏羚羊正在雪线附近奔跑;在东北的原始森林里,东北虎悄无声息地穿行。中国幅员辽阔,从海南岛到黑龙江,从东海之滨到帕米尔高原,生活着数以万计的物种。仅哺乳动物就有六百多种,鸟类超过一千三百种,这还不包括数不清的昆虫、植物和微生物。
这些生命形态千差万别。大熊猫靠竹子为生,金丝猴在树冠间跳跃,扬子鳄潜伏在水中,中华鲟逆流而上。每一种生物都有自己独特的身体结构和生活方式。但一个问题始终萦绕在人们心头:这些物种从何而来?它们一直就是现在的样子吗?
很长时间里,人们相信物种是固定不变的。猫就是猫,虎就是虎,从古至今都是如此。这种观点看似合理,因为在人类有文字记录的几千年历史中,从未见过一个物种变成另一个物种。但随着人们对地球历史的认识加深,一个全新的理解逐渐浮现。
时间的深度
理解生命演化的第一个关键,是认知地球的历史长度远远超出我们日常经验。人类的记忆和历史不过几千年,然而地球的生命故事则绵延数十亿年。在如此广阔的时间尺度下,一切生命演变都变得可能。
古时候,人们认为地球很年轻,甚至与神话传说的年代相当。但随着地质学的崛起,这种看法被推翻。科学家们注意到,厚厚的岩层和连绵的山脉其实是千万年甚至上亿年变化累积的结果。例如:
如此缓慢的变化,超出了我们肉眼和短暂生命能体会的速度。岩石的风化、山川的崛起,都是时间一点点雕琢的“艺术品”。放射性同位素测年法等先进技术证明,地球已经有约46亿年历史。若将地球46亿年历史缩成一天,人类的出现不过是“最后一分钟”发生的事情。
我们对于时间的全新理解,为解释生物的演化带来了可能。如果只给生命几千年的时长,基本看不到物种之间的改变;但如果放宽到数亿年,无数微小变异就有机会一层层积累,最终塑造出千姿百态的生命世界。正如“水滴石穿”,再细微的变化,给足够长的时间,也能产生翻天覆地的差别。
地球真正的年龄大约为46亿年,生命的历史至少有38亿年。复杂生物的演化,是无数微小变化在亿万年长河中的累积。这个“时间的深度”,为丰富多彩的生命创造了广阔舞台。
时间维度之长,恰如一部长篇史诗。每一次大规模灭绝、每一次物种诞生变革,都在地层和化石中留下证据。化石,就像一张张剧照,让我们在今天回望亿万年前的“生命大戏”。
石头里的生命史
化石是沉积于岩石中的古代生命遗物,它们可能是骨骼、贝壳、牙齿,甚至一串脚印、一团粪便。在特殊条件下,这些遗留物被矿物质逐步替代,使得它们能跨越千万年,成为我们了解远古生物的珍贵“档案”。
中国拥有丰富多样的化石资源,极大地推动了人类对演化历程的认识。例举几处具有代表性的发现:
除了这些著名的化石群,其中还包含了若干具有“连接性”意义的过渡化石。例如,辽宁“热河生物群”中发现了数量众多的近鸟类恐龙,不仅有羽毛,还能看出前肢变成翅膀的发育过程。这些“半恐龙半鸟”的化石,是演化链条上的真实“中间环节”。
化石带来的启示主要有以下几点:
- 生命的多样性在不断变化,越古老的地层,化石与现代生物的差别越大。
- 生物类型并非一成不变。不同时代的化石之间存在过渡形态,比如恐龙逐步演化为鸟类,鱼类演化为四足动物。
- 灭绝是生命史的重要组成部分。比如三叶虫、菊石等曾经繁盛一时,而今却消失在历史中;但也有个别生物抵御亿万年变迁,几乎不变。
这些延续至今的“活化石”说明,演化不是朝某个固定方向推进。如果某物种适应环境且环境长期稳定,它们就可能在数亿年里变化极少。相反,同一个地层内某些化石已灭绝,另一些则成功存活并在演化之树上繁衍出全新的分支。
实际上,人类每一次在不同地层、不同地区发现新化石,都是揭开亿万年生命史的一块拼图。石头里的遗迹,是地球这部生命史诗最直接、最有说服力的证据。
两种解释的碰撞
随着化石证据的不断丰富,人类对于生命演化本质的解释也在持续演变。从最初的直观猜想,到逐步建立科学理论,这一过程既反映了认知的进步,也展现了科学探索的复杂和曲折。
物种不变论与灾变论
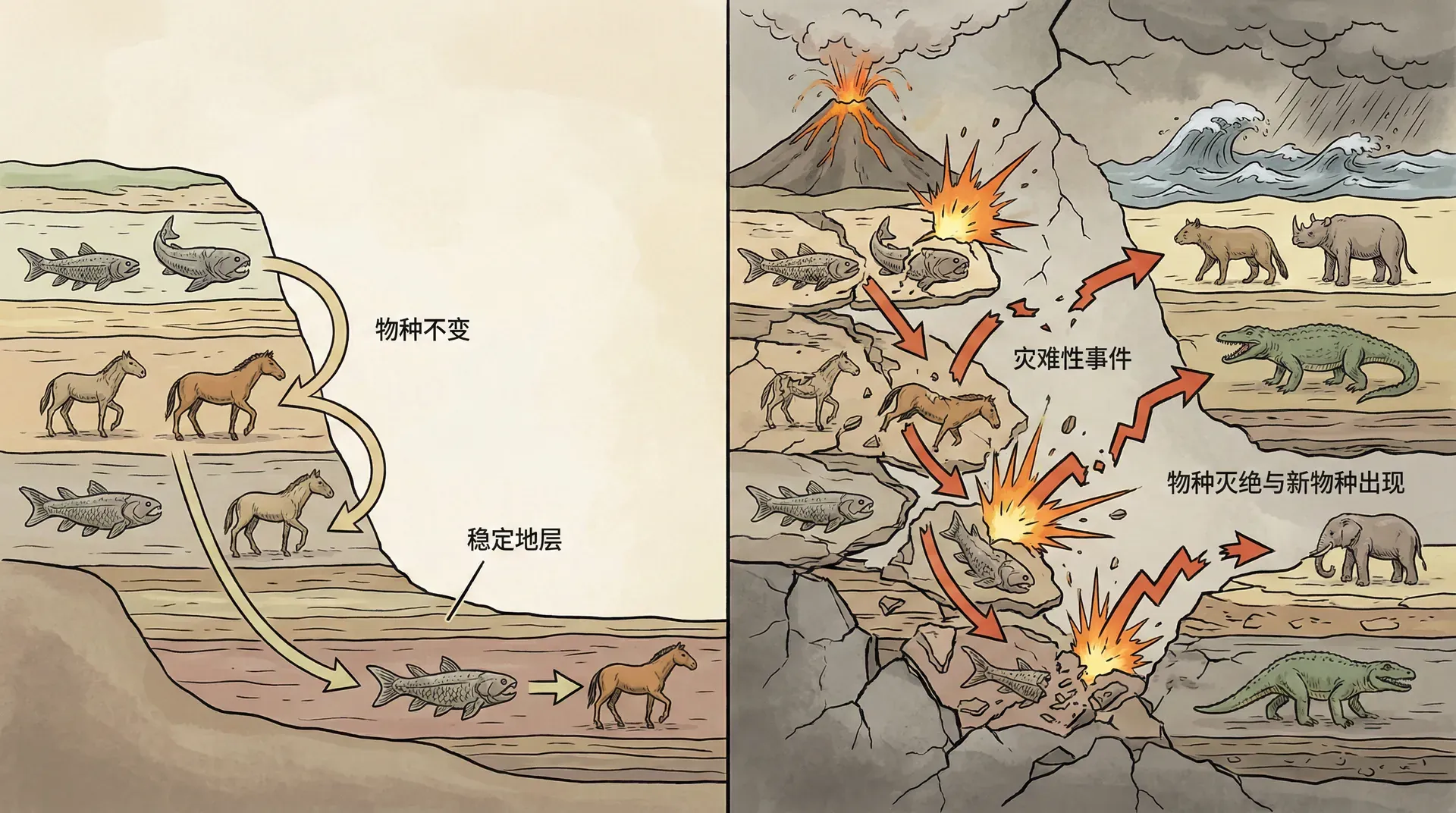
最初,在许多文明的神话和宗教框架下,人们普遍认定地球上的物种是神明在创世时一次性创造的,各自自始至终保持不变。这种“物种不变论”在历史上主宰了数千年,仿佛世界万物固若金汤。然而,化石和地层研究逐步揭示,许多物种曾经存在但已灭绝,新的生物形态又不断出现,这给物种不变论带来了极大挑战。
19世纪初,科学界流行起“灾变论”。灾变论认为,地球历史上反复发生过大规模的灾难(如洪水、火山爆发、地震等),每一次灾难都导致绝大多数物种灭绝,随后由神明或未知因素重新创造新的生命。不同地层的化石被解释为不同时代、不同灾难后出现的新生命。这一理论为化石的突然出现或消失提供了看似合理的解释,也与宗教观念形成某种妥协。
我们可以将三种早期主要假说做对比:
拉马克的“用进废退”与获得性遗传
在灾变论逐渐被证据冲击的同时,法国生物学家拉马克提出了更为“生物内部逻辑”的理论:用进废退和获得性遗传。他举例说,长颈鹿之所以脖子长,是因为祖先不断伸脖子吃高处树叶,长久下来脖子变长,后代也继承了这种性状;同理,若器官长期不用就会退化。
以下是拉马克用进废退思想的几个解释:
但遗传学的兴起很快推翻了这一设想。实验(如给老鼠多代剪尾巴)表明,后天获得的性状并不会遗传给后代。自然界多数性状的产生与积累远比“用进废退”复杂,必须涉及基因变异和选择机制,不能简单地“用则进,废则退”。
理论对比与进化论的崛起
随着达尔文与同时代科学家的探索,“自然选择”学说诞生。其核心在于:生物产生的后代远多于环境能容纳的数量,个体有各种变异,只有适应环境的变异能存活并繁殖,代代积累,有利性状逐步强化。达尔文用丰富事实论证,自然选择可以解释从简单到复杂、从古老到现代的生命全景。
三大理论的核心冲突和科学检验情况:
自然选择的提出,彻底改变了人们对生命演化的理解。在生存斗争和过量繁殖的压力下,优胜劣汰推动着群体逐渐适应环境,物种分化与多样性就此展开。生物演化,不再是静止不变的神秘过程,而是亿万年微小变化的积累结果。
科学理论的演进,是事实、实验和逻辑推理驱动下的筛选过程。每一次对生命本质的解释碰撞,都是人类理解自然的脚步更进一步。
自然的选择
每年春天,一对麻雀能孵化出四到六只小麻雀。如果所有后代都能存活并继续繁殖,几年之内麻雀的数量就会呈指数增长。但实际上麻雀的数量相对稳定。为什么?因为大部分小麻雀活不到繁殖年龄。它们会被捕食、挨饿、冻死,或者在争夺领地时战败。能够生存下来并繁殖的,只是少数。
关键问题是:哪些个体更容易存活?
答案是:那些更适应环境的个体。而适应性取决于它们身体的各种特征。
地理差异塑造的差异

东北虎和华南虎本是同一物种的不同亚种。东北虎生活在寒冷的针叶林中,体型巨大,冬季皮毛厚实而颜色较浅。华南虎生活在亚热带森林,体型较小,毛发较短且颜色更深。这些差异不是偶然的。
在寒冷的东北,体型大的虎有优势。因为体型越大,相对表面积越小,散热越少,更容易保持体温。厚实的皮毛提供额外的保温。较浅的颜色在雪地环境中更难被猎物发现。相反,在炎热的华南,体型小更有利于散热,短毛避免过热,深色条纹在浓密的亚热带森林中提供更好的伪装。
这些差异是如何形成的?在每一代老虎中,个体之间存在差异。有的体型稍大,有的稍小;有的毛发稍厚,有的稍薄;有的颜色稍深,有的稍浅。在东北的寒冷环境中,那些碰巧体型较大、毛发较厚的个体,冬天更容易存活,狩猎效率更高,因此留下的后代也更多。经过几千代、几万代的累积,整个种群逐渐向大体型、厚毛发的方向发展。在华南,选择的方向恰恰相反。
这张图展示了在寒冷环境中,拥有有利变异(厚毛)的个体数量逐代增加,而不利变异(薄毛)的个体逐渐减少。这就是自然选择的过程。
高原上的适应
青藏高原是检验生物适应能力的极端环境。那里海拔高、氧气稀薄、温度低、紫外线强。但藏羚羊、牦牛、高原鼠兔等动物在那里繁衍生息。
藏羚羊的血液有特殊之处。它们的血红蛋白对氧气的亲和力更强,能在低氧环境中更有效地运输氧气。它们的心脏和肺也比平原上的羚羊更大,能够输送更多血液。皮毛极其厚密,能抵御零下三四十度的严寒。
这些特征不是藏羚羊主动选择的结果,而是自然选择的产物。当羚羊的祖先开始进入高原地区时,大部分个体无法适应缺氧环境,很快死亡。但总有少数个体碰巧携带某些基因变异,使它们的血红蛋白效率稍高一点,或者心肺功能稍强一点。这些个体更容易存活,留下更多后代。它们的后代继承了这些有利变异,在高原上继续繁衍。经过无数代的积累,形成了今天高度特化的高原物种。
特化的代价与收获
作为食肉目动物,大熊猫的祖先和其他熊一样,是杂食性的。但大熊猫几乎完全依赖竹子为生。为了适应这种特殊的食性,它的身体发生了许多改变。
最显著的是所谓的“伪拇指”。大熊猫的前掌上,除了五根正常的指头,还有一根突出的腕骨,形成了第六根“手指”。这根伪拇指与其他手指配合,能够紧紧抓握竹竿,方便取食。这不是什么神秘的设计,而是自然选择作用的结果。在大熊猫祖先转向以竹为食的过程中,那些腕骨碰巧稍大一些的个体,抓握竹子更方便,取食效率更高,因此留下更多后代。经过漫长的积累,形成了今天这个独特的结构。
但特化也意味着丧失灵活性。大熊猫的牙齿和消化系统仍然保留着食肉动物的特征,消化竹子的效率很低。它们每天要花十几个小时进食,摄入大量竹子才能满足能量需求。这种高度特化的生活方式让大熊猫在竹林中如鱼得水,但也让它们对环境变化极其敏感。一旦竹林消失,它们很难转向其他食物。
人工选择的启示
自然选择的原理,在人类的驯化实践中得到了生动的印证。
中国人驯化家猪已有八千多年历史,野猪体型精瘦,鬃毛长而硬,性情凶猛。但人类选择性地繁殖那些体型较大、性情较温顺、产仔较多的个体,经过几百代的人工选择,培育出了各种家猪品种。太湖猪体型巨大,产仔数多;金华猪肉质细腻;荣昌猪适应性强。这些品种之间的差异,远超过野猪种群内部的差异,但它们在生物学上仍然属于同一物种。
所有的金鱼品种——狮头、水泡眼、蝶尾、珍珠鳞——都是从普通的鲫鱼培育而来。人类选择那些颜色鲜艳、形态奇特的个体进行繁殖,经过一千多年的累积,创造出了令人眼花缭乱的观赏品种。这些品种之间的外观差异巨大,但遗传上的差距其实很小,它们仍然可以相互杂交。
人工选择和自然选择的原理完全相同,只是选择的主体不同。在自然界中,环境扮演选择者的角色,决定哪些个体能够生存繁殖。在人工驯化中,人类扮演选择者的角色,决定哪些个体可以繁殖。无论哪种情况,基本机制都是:变异提供原材料,选择决定方向,时间累积造就巨变。
演化留下的痕迹
如果物种确实是演化而来,而不是各自独立创造的,那么我们应该能在生物身上找到演化历史的痕迹。事实正是如此。
用不上的器官
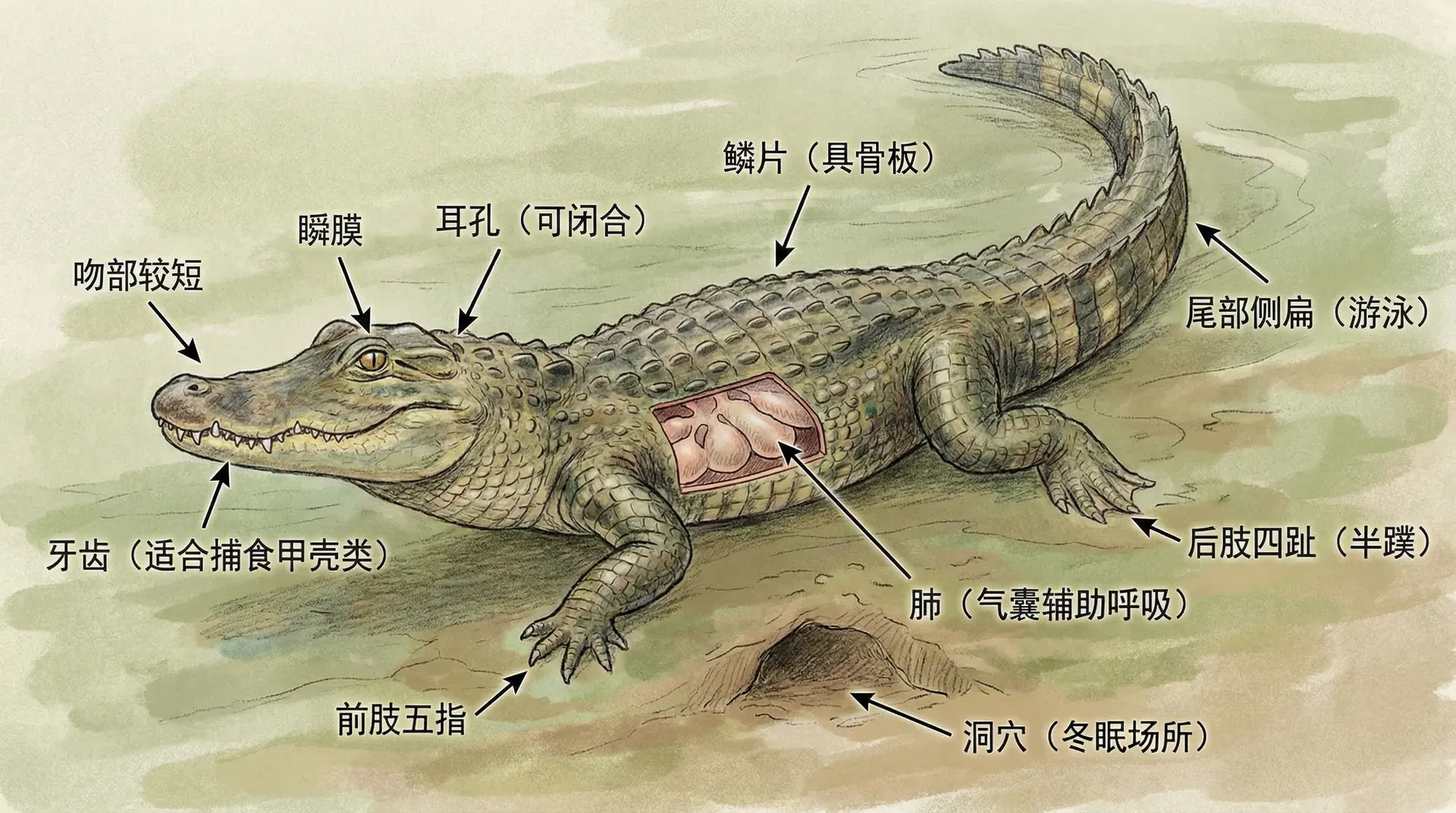
扬子鳄是中国特有的珍稀物种,也是鳄类中分布最北的种类。它的身体结构中,有一些看似没用的部分。比如,扬子鳄的后肢内侧有一些退化的骨骼结构,不与其他骨骼连接,也没有任何功能。这些是什么?
对比扬子鳄的远古亲戚,答案就清楚了。鳄类的祖先生活在陆地上,有四条完整的腿用于行走。后来一些种群进入水中生活,腿部逐渐退化,身体变得更适合游泳。那些退化的骨骼,就是祖先完整后肢的残余,就像演化历史的脚印,保留在现代鳄鱼的身体里。
中华鲟也有类似的情况。它的脊椎结构非常原始,介于软骨和硬骨之间。这种中间状态在功能上并无特别优势,但它反映了鱼类演化的早期阶段。中华鲟是所谓的“活化石”,保留了许多古老特征,让我们能够窥见几亿年前鱼类的模样。
这些痕迹器官无法用“完美设计”来解释。如果每个物种都是独立产生的,为什么要保留这些无用的结构?但如果物种是演化而来,这些结构就非常合理:它们是祖先传下来的遗产,虽然已经失去原有功能,但演化是渐进的过程,不会一下子把所有痕迹清除干净。
地理分布的启示
台湾海峡宽约130公里。在地质历史上,海平面曾经多次升降。海平面低时,台湾与大陆相连,生物可以自由往来。海平面高时,海峡形成屏障,两岸生物被隔离。
这种周期性的隔离与联通,在两岸的生物分布上留下了清晰的印记。大型哺乳动物如虎、豹、熊,历史上在台湾和大陆都有分布,因为它们能在海峡消失时迁徙过去。但许多小型动物和植物,台湾的亚种与大陆略有不同。台湾蓝鹊与大陆的蓝鹊羽色有细微差异;台湾猕猴比大陆猕猴体型稍小。这些差异是隔离期间独立演化的结果。
横断山脉这片区域地形复杂,高山深谷纵横交错,形成了无数天然的地理屏障。结果,横断山区成为生物多样性最丰富的地区之一。许多物种的分布范围仅限于某一条山谷或某一个山系,与相邻地区的近缘物种非常相似,但又有明显差异。这种格局只能用地理隔离和独立演化来解释。
相似性的深层含义
所有猫科动物——无论是东北虎、云豹还是野猫——都有相似的身体结构。它们都有锋利的爪子可以伸缩,犬齿特别发达,眼睛正对前方提供立体视觉,身体柔软适合潜行。这种相似性远远超出了功能需要。为什么不同的猫科动物不能用不同的方式来捕猎?为什么它们的相似性如此系统和全面?
答案是:因为它们有共同的祖先。所有猫科动物都继承了祖先的基本身体构架,然后在此基础上各自发展。相似性反映了共同起源,差异性反映了分化演化。
这种模式在整个生物界都能看到。所有哺乳动物都给幼崽哺乳,都有毛发,都是温血动物。所有鸟类都有羽毛,都产卵,都有相似的骨骼结构。这种成系统的相似性,最合理的解释就是共同的演化起源。
渐变的真相
要深入理解演化,就必须认识到一个核心事实:物种之间的界限并不像我们想象中那样泾渭分明。很多人通常将“物种”视为明确划分的分类单元,仿佛每个生物都自带标签,彼此之间完全隔离,界线清晰。但现实世界远比这复杂,物种分界实际上更像光谱中的色彩渐变——颜色不断过渡,没有哪一刻可以被确切标记为“分界线”。
在生物分类实践中,科学家们经常面临许多模糊和棘手的案例。比如,判断两个种群是否属于同一物种,常涉及以下情形:
这种模糊性,说明物种的形成是个连续且动态的过程。例如,东北虎和华南虎虽然目前可以杂交并且后代有生育能力,因此被归为同一物种下的两个亚种,但如果它们长期地理隔离,积累了更多差异,终有一天会变得无法杂交,这时就会被界定为两个“物种”。但究竟在何时可以“正式分家”,没有人能画出清晰界线。这一切,就像蓝色逐渐过渡到深蓝,没有哪一刻能分割两种颜色。
环状种群现象则更为特殊。例如欧亚鸫鹀(某些鸟类)环绕北极分布,各相邻地区种群可以彼此交配,但最南端两端群体却无法杂交,构成一条“物种连续体”。另外在某些昆虫和蝴蝶中,大范围分布的种群东西两端已无法交配,但通过居中地区的一系列中间型个体,连接成“环链”。
现实中的生物多样性往往打破简单的人为分类。例如:
- 蒲公英具有特殊的无性繁殖能力,经常形成成百上千个“微型物种”,划分标准极为复杂。
- 狮与虎在野外属于分布隔离的不同物种,但人工条件下可以杂交,诞生狮虎兽或虎狮兽,进一步显示边界的模糊。
- 橡树等植物属内的多个种类,尽管形态上有差别,但天然杂交现象十分普遍,很难精确区分。
人类对于种群、肤色、地理分布等的分类,其实在生物学意义上都是主观且多为表象。随着了解加深,科学家们愈发认识到:物种并不是绝对孤立、开关分明的“单元”,而是不断分化、动态变化的连续体,彼此之间由大量中间型个体和渐进过渡相连,就像树枝的分叉逐步展开。
演化论的深刻洞见在于: 生命不是固定不动的展馆,而是涌动不息的河流。我们眼中的物种,只是这股生命长河在某一时刻的一个横截面。追溯上游,可以发现所有生命最终汇聚于共同源头;而展望下游,现在的物种仍在继续演化,终将分化为全新的生命形态。正因生命本质是流动和渐变的,我们才得以目睹大自然中丰富的历史、多样的分化,以及演化背后的真实逻辑。
总结
演化论不仅是研究过去的理论,更是理解生命本质的核心框架。它揭示了生物适应性与多样性的来源:无数代自然选择下,偶发变异不断筛选,造就了大熊猫的伪拇指、藏羚羊的血红蛋白、扬子鳄的水栖特征等“适应”,也留下了消化系统、智齿等“不完美”的痕迹——这些都不是完美设计,而是遗传和环境作用下的渐进积累和妥协。
物种之间的系统相似,源于共同祖先并经历分化,构成了庞大的“生命之树”;每一物种都是漫长演化的独特结晶,失去后无法复制,因此保护生物多样性尤为重要。
一百多年来,演化论经受住了各领域无数证据的检验,是理解生命科学不可或缺的思想基石。正如生物学家所说:“脱离演化的视角,生命的一切将无从理解。”我们和大自然中的每一种生命,都是同一生命之树上的枝条,共享同一个地球与演化的历史。