遗传的复杂性与进化的随机性
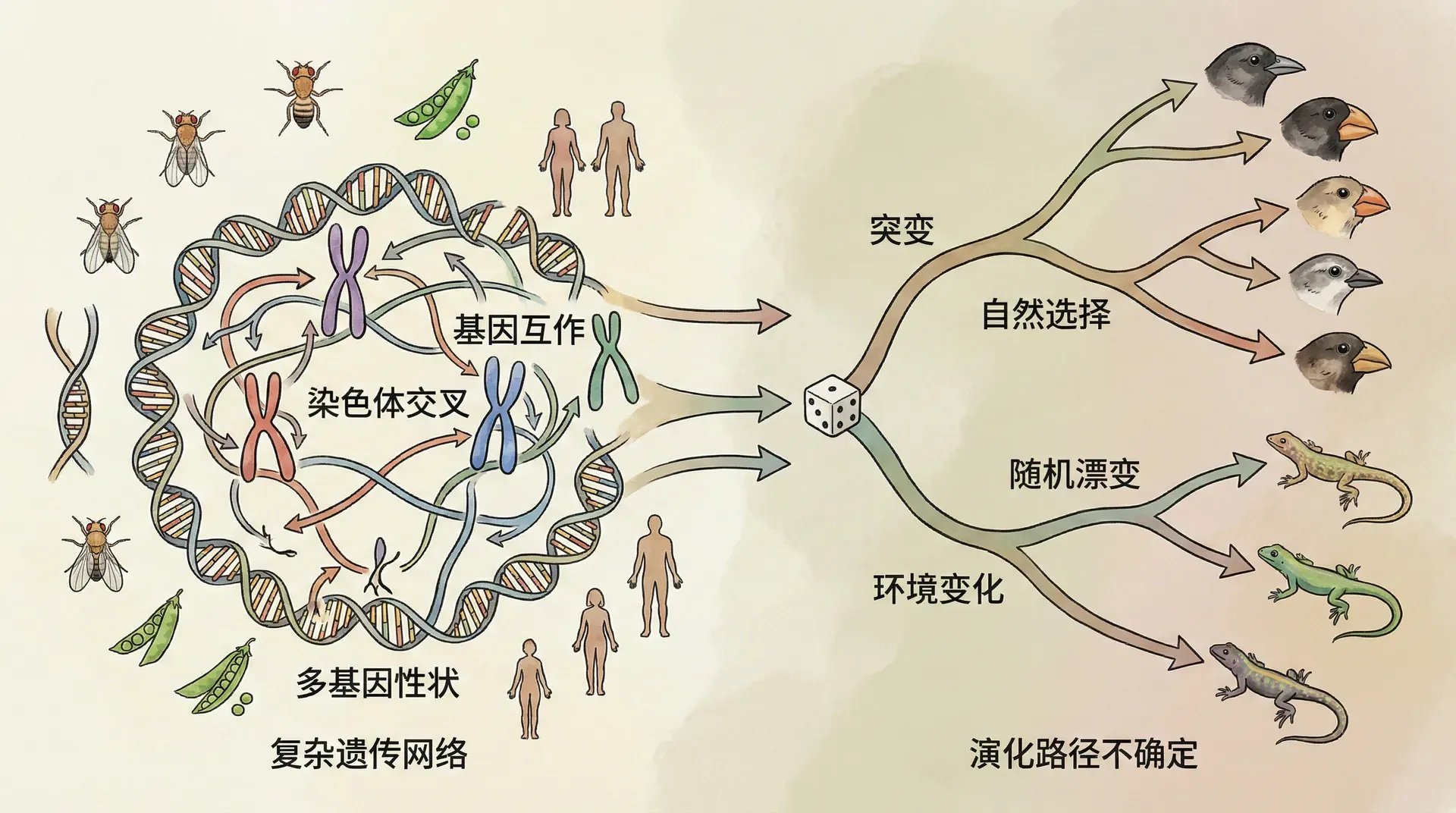
许多人下意识地以为,遗传就像是数学中的“平均值”:父母个子高,孩子也会很高;父母聪明,孩子自然也聪明。仿佛每一项性状都能直接加权继承父母的特征。然而,遗传的实际规律却远比简单的“平均”复杂得多。在生命的基因层面,性状的传递涉及到无数基因的微妙组合、显性与隐性的交互、基因突变的插入、环境与基因的共同作用等诸多因素。
正因为遗传的复杂性和随机性,才使每一个生命都是独一无二的,哪怕同胞兄弟姐妹,其表型与性格也可以有很大的差异。这种复杂性的存在,为生命的进化创造了广阔空间,也为生物多样性的产生提供了源源不断的动力。正是如此,看似“小概率”的基因重组和突变,才有机会催生出全新的特征,推动着生物世界不断演化和丰富。
不完全显性
在孟德尔的豌豆实验中,我们看到了遗传中“显性”和“隐性”性状的清晰分界。比如高茎豌豆和矮茎豌豆杂交时,第一代(F1)全部表现为高茎,矮茎性状完全没有显现,似乎“显性”基因可以彻底掩盖“隐性”基因的作用。但事实上,这种“非黑即白”的遗传方式只是众多遗传模式中的一种,自然界广阔丰富,许多遗传现象比这要复杂得多。
例如,红花金鱼草与白花金鱼草杂交以后,子代(F1)并不会全部呈现红花或白花,更不会混合成一种新颜色,而是出现了介于红与白之间的粉红色花朵。这种现象即被称为“不完全显性”,也叫“部分显性”,与孟德尔描述的完全显性大为不同。在此模式下,红花和白花的基因在杂合个体(RW)中都对表型有贡献,既没有哪一个基因完全主宰,也不是简单的“混合”,而是在性状上体现出一种中间状态。
这种不完全显性的一个有趣特点是:如果让粉色金鱼草(基因型为RW)自交或彼此授粉,所得后代表型将根据基因重组的概率重新组合,出现红花(RR)、粉花(RW)、白花(WW)三类。它们的比例大约为1:2:1,也就是红花占1/4,粉花占1/2,白花占1/4。这一比例说明,虽然杂合个体在表型上是中间的,但基因本身并未融合,每种基因型依然可以独立遗传。我们可以用下表直观表示它们的关系:
不完全显性的现象并不仅仅体现在植物身上,也可以在动物的遗传中发现。比如安哥拉兔的毛色、某些牛的毛色等,都有类似的中间表现型。实际上,不完全显性和完全显性只是遗传规律的两端,在自然界各种复杂的基因相互作用的背景下,二者之间还有许多过渡类型和其它更多变的表达方式。
不完全显性的遗传规律提醒我们,基因的表现作用并不是绝对的“有”或“无”,有些情况下基因在杂合体里会产生中间性状。这种遗传机制,大大丰富了生物性状表现的多样性,也为后续的进化和适应提供了更多可能性。在观察和研究遗传现象时,我们需要放下“简单线性”的期待,去理解生命奥秘的丰富层次。
人类血型
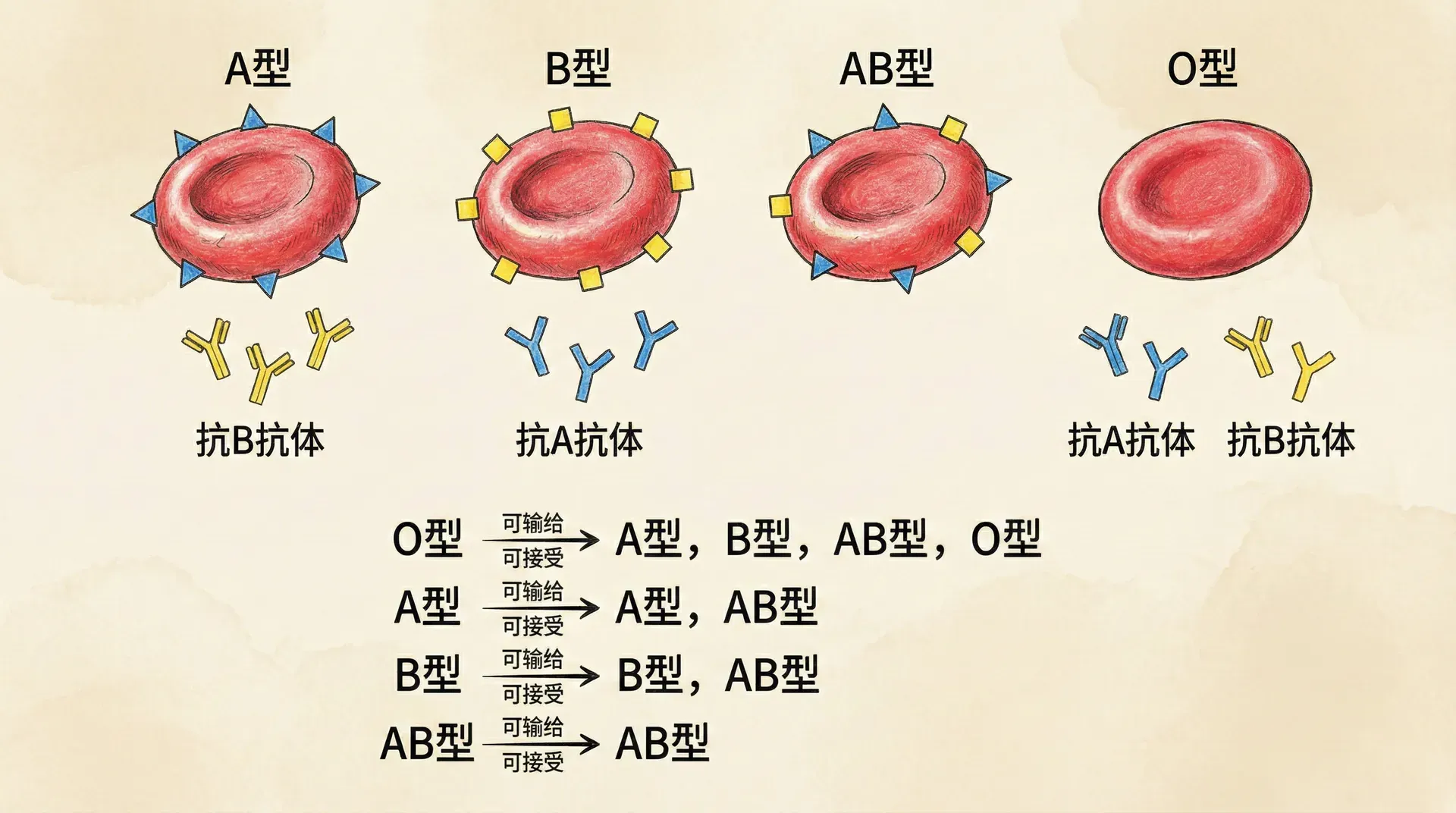
人类的ABO血型系统是理解复杂遗传模式的一个经典而生动的范例。与孟德尔豌豆实验中的单基因、两种等位基因(如高茎/矮茎)情况不同,ABO血型系统由三个等位基因(A、B和O)共同控制。这就意味着每个人体内有两份等位基因(分别来自父母),所有可能的组合有六种:AA、AO、BB、BO、AB和OO。这一差异使得血型遗传远比“显/隐”二分法复杂,并生动展现了生物遗传的多样性和规律性。
具体来说:
- 基因型AA或AO的人都表现为A型血,因为A基因对O基因显性,O的作用被完全掩盖。
- BB或BO基因型的人是B型血,同理,B对O显性。
- OO基因型才表现为O型血,这时体内既无A也无B等位基因。
- AB基因型的人表现为AB型血,这是一种“共显性”现象,A、B两种等位基因都会表达,因而红细胞表面同时拥有A、B两种抗原。
在中国人群中,ABO血型的分布还呈现出一定的地域特征。根据全国无偿献血数据,O型血约占34%,A型约28%,B型约32%,AB型相对稀少,大约6%。这些分布特点与人类历史上的迁徙、地理隔离、自然选择等诸多因素有关。例如,某些地区的O型血比例明显高于全国平均,有学者推测这可能与历史上的大规模移民、或局部人群在流行病中的生存适应有关。
血型遗传的规律十分明确和可预测,是中学生物课本常见的推理题型之一。A基因和B基因均对O基因显性,但A、B之间并不是互相“压制”,而是“并列表达”,也就是共显性。因此,父母血型的组合直接决定了他们子女所有可能的血型范围(但不一定等概率出现)。
这些组合可以用“潘尼特方格”来推演所有基因型的组合可能,也帮助人们理解何时可能出现某种“罕见”血型。从表格我们看到,只有A型和B型父母组合时,孩子可能拥有A、B、AB、O四种血型的任意一种。而O型父母则只能生出O型孩子,无论多少代都无法自然产生出A型、B型或AB型后代。
血型遗传具有明确规律,因此常用于亲子关系排除。在法医学和医疗中,如果孩子血型与父母血型组合无法通过遗传规律解释,可以初步排除亲缘关系。例如,两个O型父母不可能有AB型孩子。不过,血型主要适用于“排除”,不能单独作为确定依据,因为多种父母血型组合可能产生相同血型的孩子。
此外,ABO血型只是众多血型系统中的一种。在临床输血和移植时,常常还需考虑Rh血型系统等其他抗原。但ABO血型由于拥有清晰的遗传逻辑,成为遗传学教学和日常生活中最具代表性和实用意义的一类。有趣的是,研究还发现某些血型与疾病易感性、感染风险等存在一定关联性,这说明血型不仅仅影响输血匹配,也可能与人体健康有一定关系,显示出血型研究的更宽广价值。
因此,ABO血型给我们展示了一种复杂但规律清晰的非单纯“显/隐性”遗传模式,也让我们认识到,现实世界的基因遗传比课本示例更加丰富和耐人寻味。
多基因控制的性状
在生物遗传中,许多常见且重要的性状并非由单一基因所控制,而是受到多个基因(通常称为多基因或数量基因)的共同作用。这类性状被称为“多基因控制性状”或“数量性状”。与孟德尔经典豌豆实验中“紫花与白花”、“高茎和矮茎”的两极分化不同,多基因性状通常表现为连续的表型变化,而不是简单的“非此即彼”分布。
让我们来看一些例子:
水稻株高
水稻的株高性状,并不是由某一个“高秆基因”或“矮秆基因”决定的。研究显示,至少有十几个不同的基因座位共同影响株高。每一个基因位点的贡献大小不同,有的影响显著、有的微乎其微,这种累加效应导致水稻的株高呈现出从非常矮到非常高的连续分布,中间过渡类型极为丰富,远非“高-矮”两极能概括。
上世纪六十年代,袁隆平院士发明杂交水稻技术时,利用了多基因控制株高和产量的科学原理。科学家通过繁杂的选育和反复杂交,把有利的株高相关基因组合在一起,既实现了增产,又让植株不易倒伏,大大推动了粮食生产。
人类身高
人类身高是经典的多基因性状之一。最新的基因组研究表明,决定身高的相关基因位点多达数百个甚至更多,每个基因的单独效应都很小,只有在整体叠加下才形成所见的身高差异。因此:
- 父母身高高,子女身高通常也高,但后代未必完全复制父母的具体数值,因为每次遗传、基因重组带来组合的新可能;
- 矮个父母同样可能生出较高的孩子,归因于对方家族遗传、基因组合偶然等原因。
影响身高的部分因素
在中国,历史以来南北方人群的平均身高存在差异。一般北方人稍高,南方人偏矮一些,但这并不是因为“北方有特殊高个基因”,而主要是整个种群中多种微效基因频率及营养、生活习惯长期综合作用的结果。事实上,多基因性状对环境的敏感度比单基因性状强烈得多。
多基因性状与环境的关系
多基因控制的性状还有一个显著特征:环境的影响往往非常明显,甚至可以与遗传因素分庭抗礼。比如单基因疾病“生/不生”往往很绝对,但身高、体重、作物产量等,多数情况下是遗传与环境的叠加。
例如,遗传上有“高个”潜质的孩子,如果成长过程中长期营养不良,最终身高也会大打折扣。反之,中等遗传潜力的个体若获得良好的饮食、卫生和运动,也有机会接近其种群的上限。这种现象在多基因性状中最为常见。
综合来看,多基因控制的性状是遗传学研究与实际育种、健康管理中尤为重要的对象。只有在全面理解数量基因与环境相互作用下,才能科学解释和合理调控这类性状的变异与改良。
基因与环境的交响曲
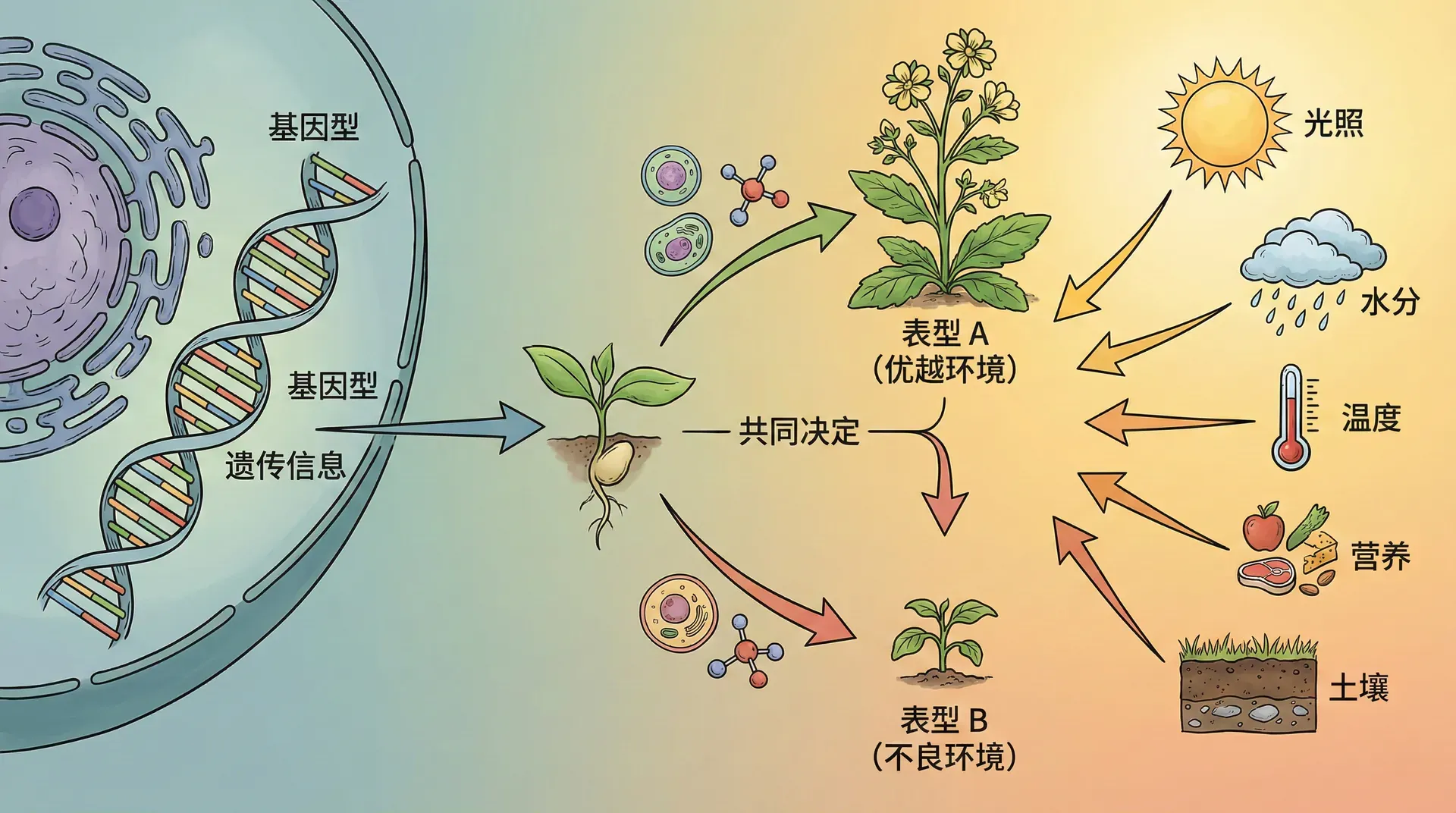
基因不是在真空中发挥作用的。基因表达需要特定的内环境和外环境条件,而环境因素的变化可以显著改变基因的表达效果。这种基因与环境的互动关系,在人类适应不同生存环境的过程中体现得尤为明显。
生活在青藏高原的藏族人群为我们提供了一个绝佳的例子。高原地区氧气稀薄,大气压只有平原地区的60%左右。在这种环境下,普通人会出现高原反应,包括头痛、呼吸困难、心跳加速等症状。但世代生活在高原的藏族人群却能自如地在这种环境中生活劳作。
研究发现,藏族人群在某些基因上发生了特殊的变异。其中最重要的是EPAS1基因(有时也称为“高原适应基因”),这个基因影响着身体对低氧环境的反应。藏族人群中这个基因的特殊版本帮助他们在低氧环境下维持正常的血红蛋白浓度,避免血液过度黏稠。有趣的是,如果具有这种基因变异的藏族人长期生活在平原地区,这个基因的"优势"就不再明显,甚至可能变得无关紧要。
基因型是生物体的遗传蓝图,但表型是基因与环境共同作用的结果。同样的基因在不同环境中可能产生不同的表现,这就是基因表达的环境依赖性。
过去几十年里,中国青少年的平均身高显著增长。研究显示,19岁中国男性的平均身高已达到约 175.7 cm、女性约 163.5 cm,与上世纪八十年代相比增长明显,这种变化速度远远超过基因突变和自然选择可能在短期内产生的遗传效应,主要反映了营养改善、卫生条件提高和生活水平提升等环境因素的影响。
真正的原因在于营养条件的显著改善。蛋白质、维生素和矿物质的充足摄入为生长发育提供了物质基础。一个人可能遗传了“高个子”的基因组合,但如果在生长发育期长期营养不良,最终身高可能远低于基因潜力。反之,即使遗传潜力一般,良好的营养也能让身高接近或达到遗传上限。
这个现象说明,我们观察到的生物性状(表型)是基因型与环境共同作用的产物。对于多基因控制的性状尤其如此,环境因素可以在相当大的范围内调节基因的表达效果。
进化的随机性
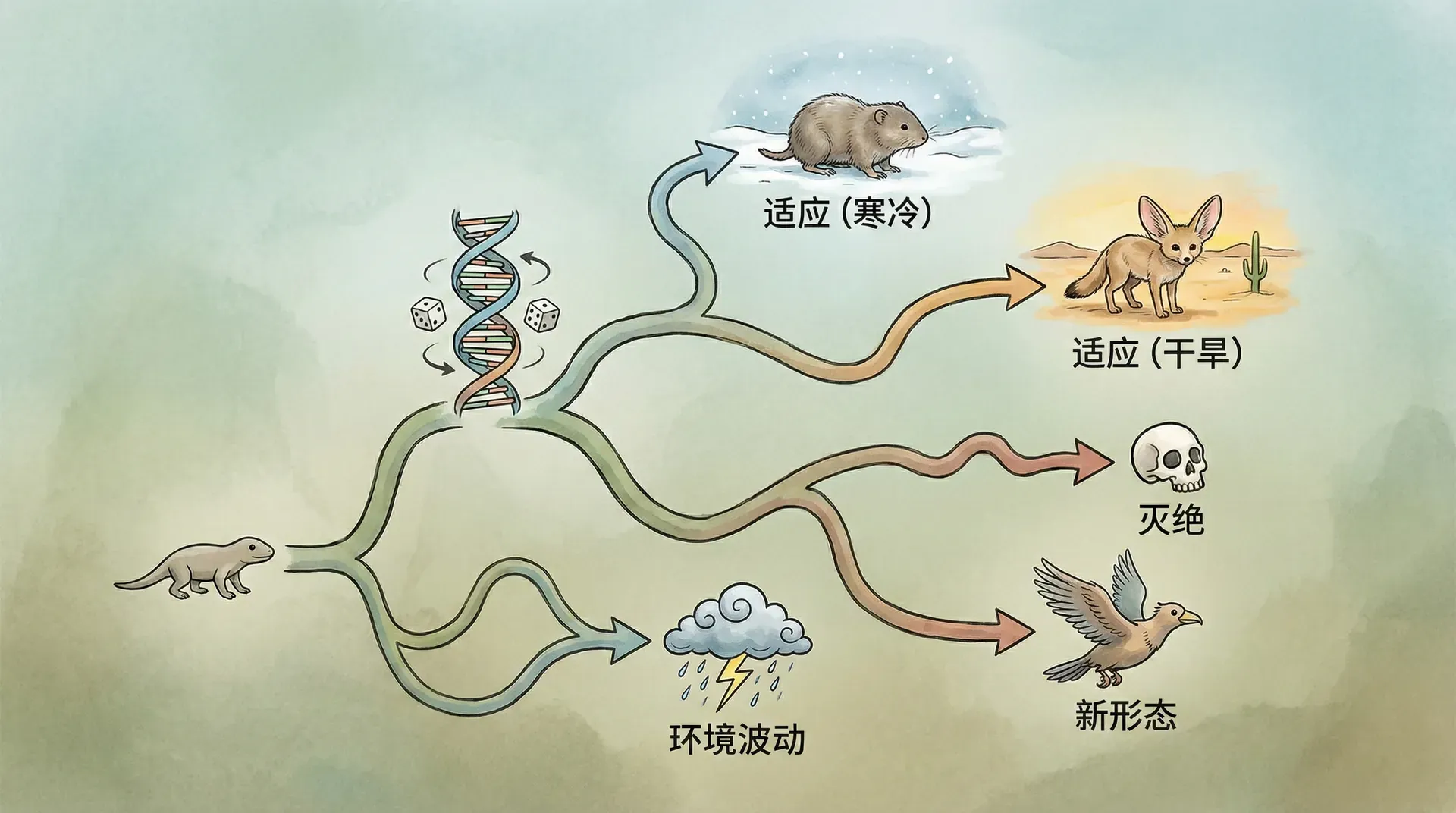
当我们理解了遗传的复杂性之后,一个更深层的问题浮现出来:生物是如何演化的?从简单的单细胞生物到复杂的多细胞生物,从海洋生物到陆地生物,从爬行动物到哺乳动物,这个漫长的过程是否有某种“方向”或“目的”?
答案可能让人意外:进化本质上是一个随机过程。
基因在世代传递中会发生突变。这种突变就像是遗传密码的“打字错误”。DNA复制是一个极其精确的过程,但每复制数亿个碱基对,就可能出现一次错误。大多数突变是中性的,既不明显有利也不明显有害。少数突变有害,极少数突变恰好在特定环境下有利。
关键在于,突变的发生是完全随机的。细胞不会“知道”什么样的突变是有用的,不会根据环境需要来定向产生某种突变。一个生活在寒冷环境中的动物不会因为需要保暖而“主动”产生增加毛发的基因突变。突变就像是掷骰子,每一次都是随机的,与需求无关。
但这不意味着进化完全是盲目的碰运气。随机突变提供了“原材料”,而自然选择则起到了“筛选”的作用。有害的突变会被淘汰,因为携带这些突变的个体生存和繁殖能力下降。有利的突变会被保留和扩散,因为携带这些突变的个体更容易生存和繁殖。中性突变则随着基因的随机组合在种群中漂移。
这个机制的精妙之处在于:虽然突变是随机的,但选择是定向的。随机过程加上非随机的筛选,最终导致了生物适应环境的表象。
地理隔离
物种的形成往往始于地理隔离。当一个原本统一的种群被自然屏障(如山脉、河流、海洋)分隔成相互隔绝的小种群后,每个小种群会经历独立的演化历程。
大熊猫为我们提供了一个生动的案例。目前中国的野生大熊猫主要分布在两个地理区域:一个是秦岭山脉,另一个是四川的岷山、邛崃山、大相岭等山系。秦岭种群与四川种群之间被平原和丘陵地带隔开,基因交流极其稀少,可以视为两个相对独立的种群。
在漫长的隔离过程中,两个种群各自经历着基因突变和随机组合。某个突变可能在秦岭种群中出现,通过繁殖逐渐扩散到整个秦岭种群,但永远无法传播到四川种群。同样,四川种群中出现的新突变也无法传递给秦岭种群。
随着时间推移,两个种群在基因组成上会出现越来越多的差异。这种差异的积累不仅来自新突变,也来自基因频率的随机漂变。在小种群中,即使某个基因变异本身没有明显的优势或劣势,其频率也可能因为繁殖的随机性而发生波动。几代之后,某个等位基因可能在一个种群中变得更常见,而在另一个种群中变得更稀少,甚至完全消失。
下面的图表模拟了地理隔离后两个大熊猫种群中某个中性基因频率的变化过程。我们可以看到,即使在没有自然选择压力的情况下,仅仅因为随机的繁殖过程,两个种群的基因频率也会逐渐分道扬镳。
这张图表展示了一个重要的进化学原理:地理隔离的种群会因为随机因素而在遗传组成上发生分化。初始时刻,两个种群中某个基因的频率都是50%。但随着世代更替,秦岭种群中这个基因的频率逐渐上升到80%左右,而四川种群中则下降到20%左右。这种分化并不需要自然选择的参与,纯粹是小种群中随机繁殖导致的结果。
实际观察也证实了这一点。研究人员发现,秦岭大熊猫与四川大熊猫在形态特征上存在一定差异。秦岭亚种的头骨较小,牙齿较大,胸部有褐色斑块的比例较高。这些差异虽然不足以将它们划分为不同物种,但确实反映了长期地理隔离造成的遗传分化。
自然选择
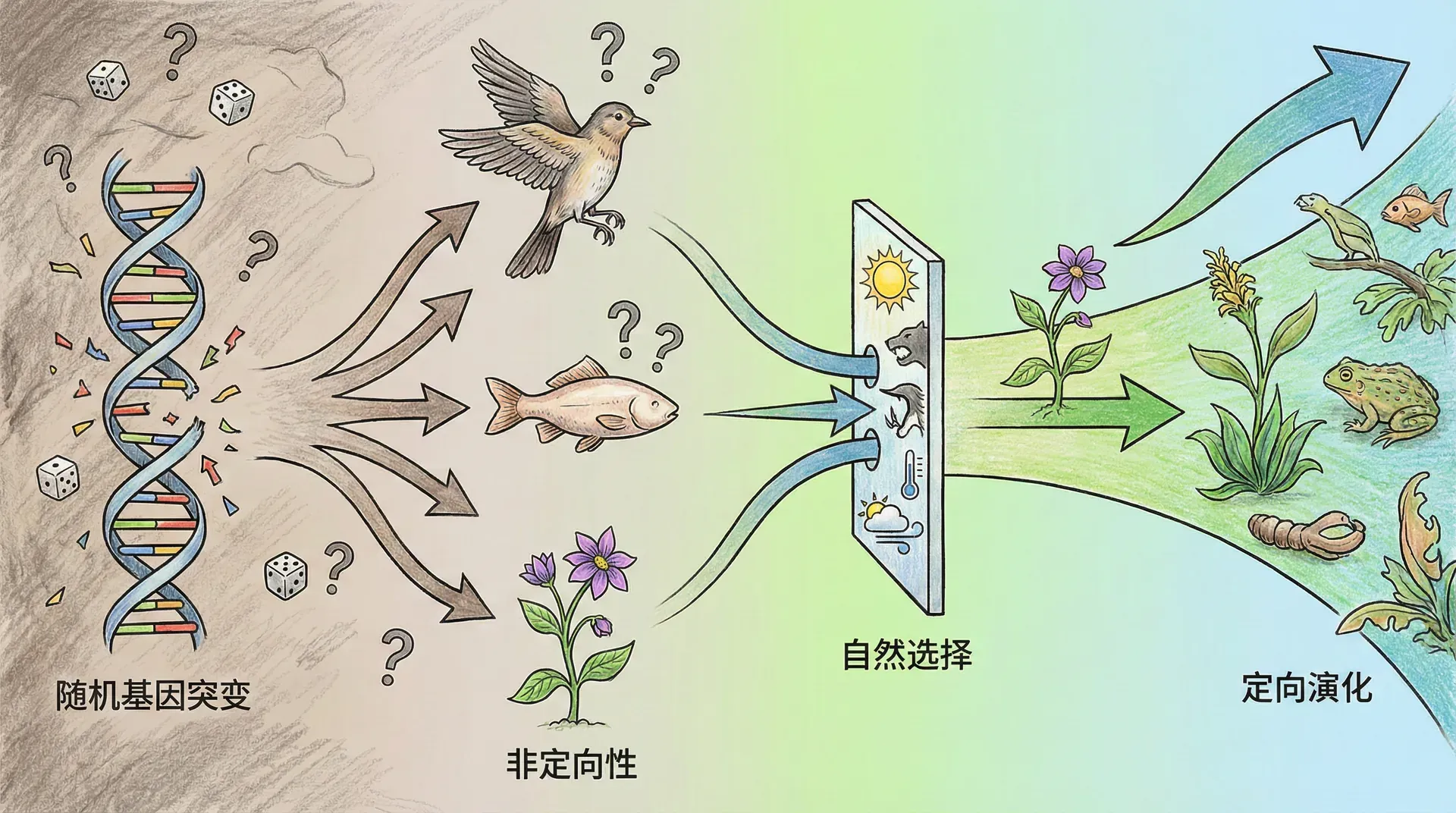
如果说基因突变是随机的,那么自然选择就是定向的。自然选择不会“创造”新基因,但它会决定哪些基因变异能够保留下来,哪些会被淘汰。
青藏高原的牦牛为我们展示了自然选择如何塑造生物的适应性特征。牦牛长期生活在海拔3000米以上的高原地区,那里不仅氧气稀薄,气温也极低。普通牛类很难在这种环境下生存,但牦牛却能在这里繁衍兴旺。
牦牛的高原适应能力并非一朝一夕形成的。在漫长的进化历程中,牦牛祖先种群中不断发生着随机的基因突变。大多数突变是中性的或有害的,但偶尔会出现一个恰好有利于高原生存的突变。比如,某个突变可能使得血红蛋白与氧气的结合能力稍微增强,或者使得毛发更加浓密保暖。
携带这些有利突变的个体在高原环境中生存能力更强,更容易活到繁殖年龄,也能养育更多后代。于是,这个有利突变在种群中的频率会代代增加。经过成千上万代的积累,多个有利突变在牦牛基因组中聚集起来,最终形成了我们今天看到的高原适应特征。
这个过程的关键在于环境的“选择”作用。在高原环境中,某些基因变异是有利的;但如果环境改变,这些变异的优势可能就不复存在。这就解释了为什么不同环境中的生物会演化出不同的特征。
自然选择是一个缓慢的过程。它需要多个世代的时间才能产生明显效果。一个有利突变从在种群中出现到成为普遍特征,可能需要几百甚至几千代。这也是为什么物种演化需要以百万年为单位的时间尺度。
随机突变提供了遗传变异的原材料,自然选择从中筛选出适应环境的变异,地理隔离则允许不同地区的种群沿着不同方向演化。这三者共同构成了进化的基本机制。没有任何神秘力量在指引进化的方向,也没有任何预设的目标。生物适应环境的精巧特征,完全可以由随机突变加上自然选择来解释。
特化
在进化过程中,许多生物会朝着某个特定方向发展,越来越适应某种特定的生活方式或环境,这种现象被称为特化。特化可以赋予生物在特定环境下的巨大优势,同时也意味着一定的代价和脆弱性。
以下是特化现象的两个例子:
上述表格总结了两种特化生物及其适应环境、获得的优势与面临的潜在代价。
特化有时还会导致某些身体结构的简化或消失。例如,在中国的岩溶洞穴中,许多洞穴鱼的祖先曾有正常的眼睛和色素,但在漫长的进化历程中,因为黑暗环境下这些结构失去了作用,不再保留下去,最终演化出无眼无色素的盲鱼。
进化不一定意味着变复杂。当简化能够提高适应性时,进化也会朝着简化方向进行。洞穴鱼类失去眼睛和色素,就是通过简化来适应环境的典型例子。
人类自身的进化足迹
人类也在经历着进化过程,虽然现代文明在很大程度上隔离了我们与自然选择的直接作用。观察人类身体结构的变化,可以看到进化的痕迹,尤其是在牙齿数量的变迁上尤为明显。
我们来看不同时期人类牙齿数量的概况:
最明显的例子就是智齿(第三磨牙)。我们的远古祖先主要以粗糙的植物性食物为主,32颗牙齿都是必需的。但自从人类学会用火和工具,饮食变得更精细,逐渐不再需要全部牙齿,颌骨也跟着变小。现代人口腔医学调查显示,中国人群中大约20-30%的人天生缺少1颗或多颗智齿,阻生智齿和拔除智齿都非常常见。这是人类简化进化的一个活生生实例。
中国的古人类化石(如北京猿人)也印证了这一趋势:50万年前的祖先颌骨更粗壮,牙齿更大更全;而现代人不仅牙齿数量减少,颌骨和牙齿体积也明显缩小。这一切与食物加工技术的进步密不可分。
总结
到20世纪初,生物学的几大核心理论逐渐汇聚,描绘出演化机制的主干轮廓:地质学证实了地球和生命历史的悠久,达尔文的自然选择解释适应性,孟德尔定律阐释了遗传规律,而突变理论揭示了变异产生的途径。这些观点共同揭示,生命的多样性是基因变异与自然选择在广阔时间尺度下交互作用的结果。没有任何先验目的或方向,生物会因随机变异产生差异,经过非随机的选择,逐渐形成适应环境的各种形态。
然而此时,科学家们对于“基因”本身的理解还非常有限:基因到底以什么物质和结构存在?它们如何实现自我复制和指导生命的发育?这些更为根本的问题,将推动遗传学进入崭新的分子时代。