生物之间的相处之道
在长江流域的稻田里,你会发现水稻根部长有小瘤状突起,里面住着能将空气中氮气转化为水稻可用氮肥的根瘤菌。水稻为根瘤菌提供有机物和庇护,根瘤菌帮助水稻获取养分,双方互利共生。这种关系在豆科植物、树根与真菌等之间也广泛存在,是生态系统的重要基础。
当然,稻田中还有很多其他关系:害虫啃食水稻叶片,杂草和水稻争阳光和养分,水中线虫寄生其他生物,青蛙捕食害虫又被蛇猎食,还有各种微生物既有竞争也有合作。在同一片稻田里,生物们有时合作共赢,有时你争我夺,也有寄生谋利,关系错综复杂。
这些关系不仅限于稻田,在森林、草原、湖泊甚至家中一花一草之间都能见到。生物间合作、竞争、寄生、掠食等多种模式,共同构成了多样且动态变化的生态系统,这正是我们本章要探讨的主题。
生物之间有哪些相处模式
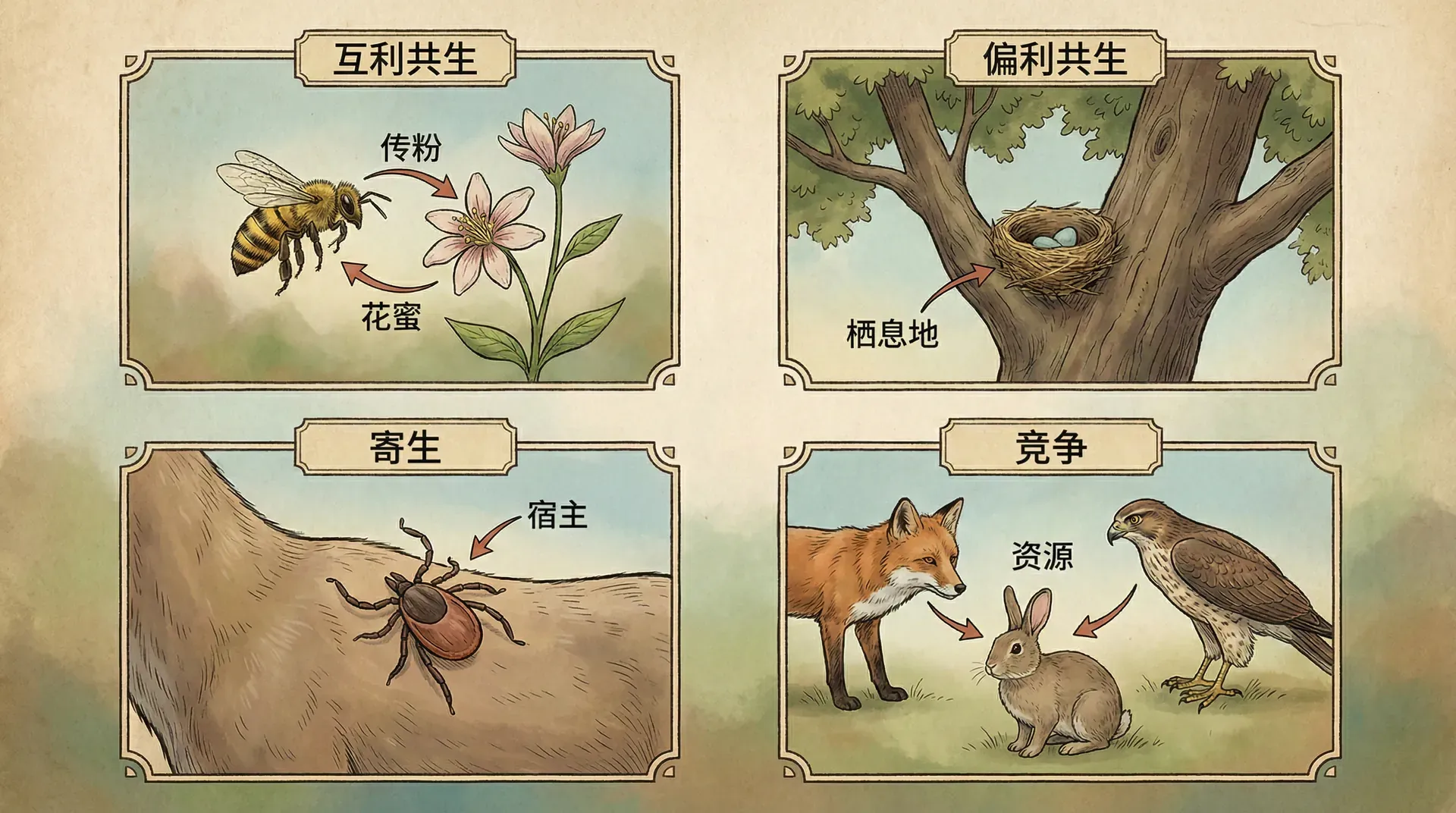
在一个热闹的菜市场,不同的人有不同的关系:有的是买卖双方,互惠互利;有的是竞争对手,争夺同样的顾客;有的是小偷和受害者,一方得利一方受损。生物世界里的关系模式,其实也是这样分类的。
我们可以用一个标准来区分这些关系:看看互动的双方,各自的生存和繁殖能力是增加了(+)、减少了(-)、还是没什么变化(0)。
在我们的日常生活中,最容易观察到的就是偏利共生关系。城市里的麻雀就是一个典型例子:它们在人类的房檐下筑巢,吃人类丢弃的食物残渣,但对人类的生活几乎没有影响。小区花园里的青苔也是如此,它们长在石头表面,获得了一个落脚的地方,但石头本身既不受益也不受害。
生物之间的关系并不是固定不变的。同一种生物在不同环境下,可能与其他物种建立完全不同的关系。比如,某些真菌在营养充足时是植物的互利伙伴,但在食物匮乏时就会变成寄生者,从植物身上掠夺养分。
寄生生活的代价
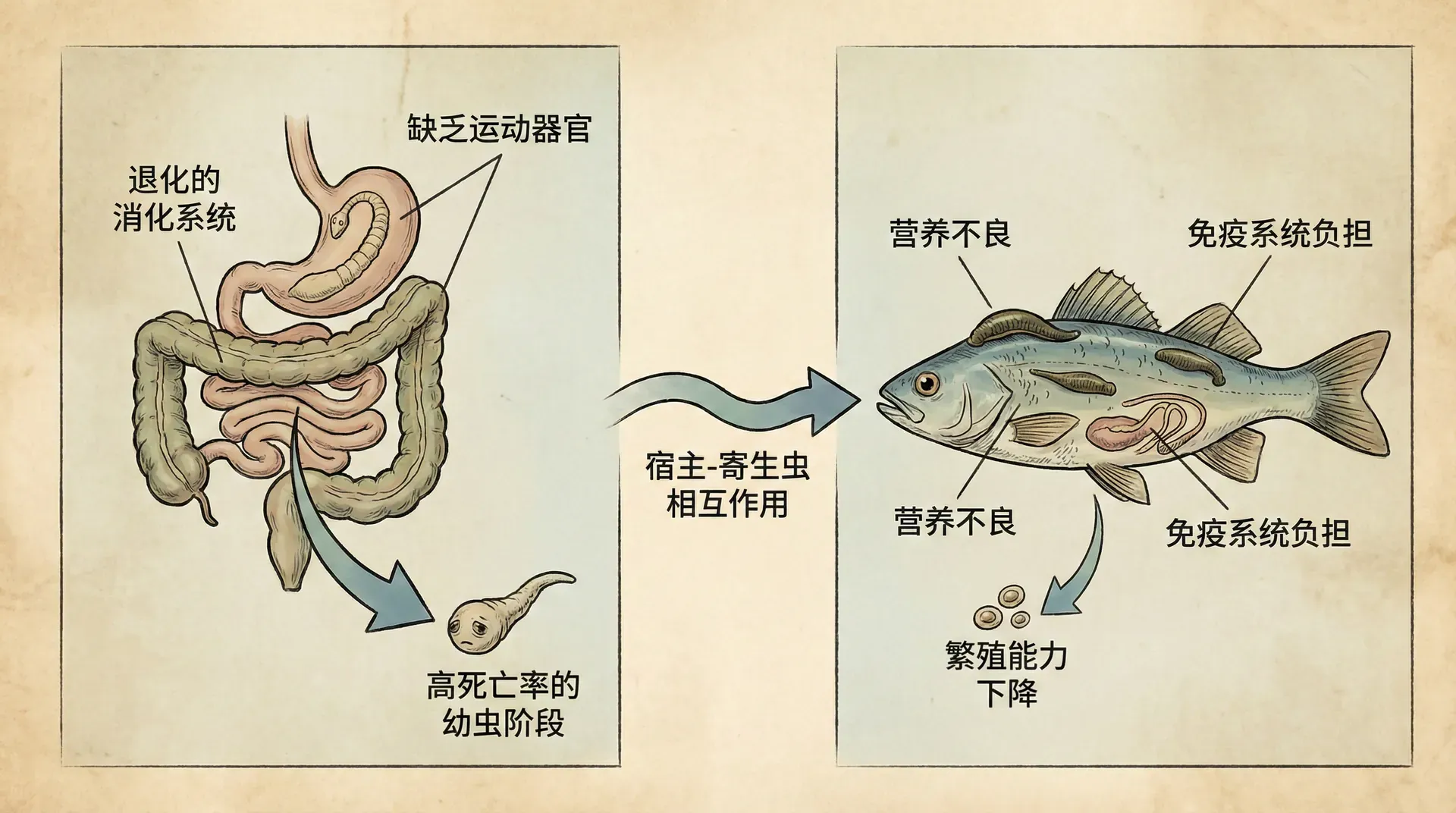
我们来聊一个让人疑惑的问题:寄生虫依靠宿主生活,为何有些寄生虫却会让宿主死亡?这样一来,寄生虫不就自断后路了吗?
长期以来,生物学家认为寄生虫应该倾向于“温和”,以便宿主活得更久,从而让自己有机会繁殖更多后代。但现实远比这一假设复杂。20世纪80年代,科学家们提出新的理论,认为寄生虫的"毒性"(即对宿主造成的损伤程度)其实是多种因素平衡的结果。简单来说,寄生虫既要提升传播效率,又要确保不至于让宿主太快死亡,失去繁殖机会。
想衡量寄生虫的成功,可以看它在宿主群体中的平均传播数——即每个感染个体平均能传染给多少健康个体。这个数字越大,寄生虫就越“成功”。而影响这个数字的关键因素有:
举例来说,如果寄生虫在宿主体内疯狂繁殖,产下大量后代,自然更容易传播出去。但这种策略往往会严重损害宿主健康,甚至导致宿主迅速死亡或痊愈后产生免疫,从而大大缩短了它的传播时间。反之,如果寄生虫太温和,虽然宿主可存活较久,但它扩散的速度又跟不上。
于是,演化出了这样一个权衡:
- 高毒性:传播快但时机短,宿主快速死亡或康复,机会稍纵即逝
- 低毒性:传播慢但时机长,宿主生存期长但传播范围有限
- 中等毒性:两者平衡,在特定环境下获得最大繁殖收益
如下图所示:
以血吸虫病为例,不同环境下寄生虫毒性的最优策略也不同。中国南方曾是血吸虫高发区,但有意思的是,人群密集和卫生条件差的地方,血吸虫毒性反而偏高。这正是因为:
- 病人即使很快重病倒下,虫卵也能轻松找到新宿主,传播不成问题;
- 于是寄生虫可以“激进”繁殖,不必太“顾及”宿主身体。
而在人烟稀少或卫生状况良好的地区,若宿主过早倒下,反而难以传播。只有那些“留情面”、让宿主保持活力的寄生虫变异能延续下去。类似的现象在家禽流感、兔出血症等动物流行病中也可观察到。
一个身体里的“竞争赛”
寄生虫并非总是独自行动。在同一宿主体内,往往存在不同“品系”甚至不同物种的寄生虫。这时会发生什么?
假如你和几个陌生人被困在粮仓。粮食总量有限,大家可以慢慢分,但也可以疯狂争抢。若有人开始抢,其他人都不得不跟,结果大家都没捞到多少好处。这就是著名的“公地悲剧”。
寄生虫之间的内部竞争,往往推动它们提高繁殖速度、增强毒性,抢在对手繁殖之前尽量利用宿主的资源。实际研究表明,多重感染时病原体的毒性确实较高。比如某些艾滋病毒感染者体内,如果出现多个病毒亚型共存,病毒复制和免疫系统损害速度都会加快。
传播方式决定毒性演化
寄生虫毒性的演化,也牢牢受到“传播方式”的制约,主要有水平传播和垂直传播两大类型:
案例说明如下:
科学家在无花果小蜂及其寄生线虫系统中做过实验证明:
- 如果无花果中只有一只雌蜂产卵,线虫只能通过雌蜂后代复制(即垂直传播),此时线虫对小蜂几乎无害;
- 若多只雌蜂在同一果实产卵,线虫可在宿主之间传播(水平传播增多),此时更容易演化出较高毒性,迅速复制抢占资源。
该实验生动展现了传播方式如何影响寄生虫与宿主关系的演变方向。
合作共赢的演化
在自然界中,寄生是单方面获利,而互利共生则意味着双方都能受益。这种合作关系非常普遍,从高山到大海、从森林到荒漠,几乎每个生态系统中都有生物“合作”的身影。
我们熟知的例子里,蜜蜂和油菜花的关系堪称典范。蜜蜂采集花蜜、花粉——这为它们提供了能量和蛋白质来源;而油菜花获得了传粉的帮助,可以更好地繁衍后代。在云南的热带雨林中,还有“蚂蚁保镖”保护合欢树,蚂蚁驱赶食叶昆虫,作为回报,植物分泌蜜露给蚂蚁吃。而在我们的肠道里,数以万亿计的细菌帮我们消化纤维素、合成维生素,而我们则提供温暖而稳定的“家园”。
我们可以用如下表格对比几个常见的互利共生案例:
但为什么在竞争激烈的自然界中,会演化出愿意向“别人”付出资源的生物?合作的关系是怎样逐步形成、变得牢不可破的呢?
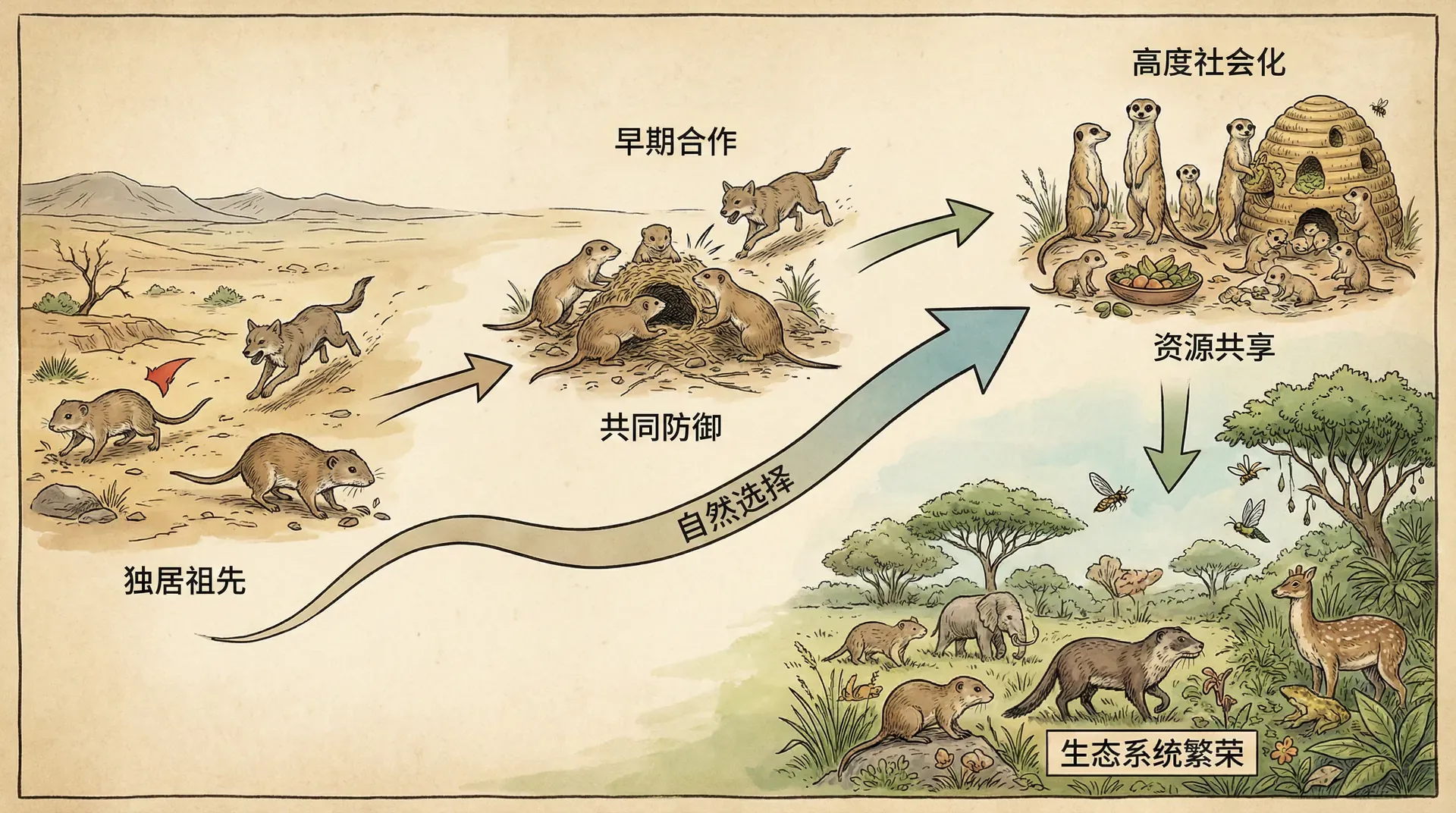
合作的起点
理论研究表明,互利共生能够建立主要依赖于两个关键因素:初始收益是否足够高、双方的空间关联度。
1. 初始收益——合作能否“起步”
假如有两种生物初次合作。比如,某植物的根分泌糖分(自己要花能量制造),吸引真菌过来。真菌帮它吸收水分和矿物。但如果植物付出的糖太少,真菌“收入”不高,就不会留在它身边,合作便终止。只有当付出的糖所吸引的真菌能让植物收获更多水分和矿物,补偿甚至超过付出时,这种合作才会被“选中”并延续下去。
这里就有一个“临界阈值”——合作只有在双方投入与回报的比例跨过某个点,才能真正启动,并且会随着演化不断优化(双方努力,付出变少、回报增多)。
2. 空间关联度——合作能否“稳定”
如果两个物种常常随机遇见,很容易“擦肩而过”,合作难以长期维持。但如果两者固定配对合作,比如栖息在彼此身上、形成家族传承,则更容易形成稳定互利的关系。
举例来说:
- 根瘤菌与豆科植物:根瘤菌从土壤进入豆科植物,定居在根瘤这个“封闭小屋”中不再外流。植物则可以通过调整给不同根瘤的养分多少,“奖勤罚懒”,只支持固氮效率高的细菌。
- 榕树与榕小蜂:每种榕树都有唯一专属的榕小蜂传粉。雌蜂钻进榕果内部,产卵又传粉,蜂的后代全在果内长大,羽化、交配、再去别的榕果。这极致绑定让合作关系异常稳固。
从剥削到共生
有趣的是,互利共生关系未必起点就是“合作”,不少合作关系的历史都从剥削、甚至“敌对”演化而来。
理论模型认为,初始时某一方进入另一方体内掠夺资源,宿主也被迫演化出各种“护身符”——比如加厚细胞壁、分泌毒素,对抗寄生者。然而,这些防御并非免费:加厚细胞壁、制造毒素都要耗费能量。如果防御产生后依然维持,但寄生者却离开,防御反倒成了包袱,让宿主竞争中败下阵来。
时间一长,“轻装上阵”的个体被淘汰,只剩能和寄生者共处的个体。这时双方虽然本质还是单向的“资源流动”,但已经彼此依赖、谁也甩不掉谁——共生关系诞生了。
我们可以列出从剥削到合作的过程主要阶段:
在现实实验中,科学家曾让变形虫吞下某种初为寄生者的细菌,最初它们使变形虫存活率下降。但若干年后,含细菌的变形虫反倒比未感染者活得更好——共生、乃至依存,悄然发生。
合作关系的演化过程告诉我们:现今和谐的“双赢”,往往源自曾经的激烈博弈和复杂历史。理解这一过程,有助于我们欣赏自然界中丰富多样、互利共存的奥秘。
如何防止背叛
互利共生关系虽然对双方都有利,但它面临一个严峻的挑战:背叛者的入侵。如果有个体能享受合作的好处却不付出代价,它就能占便宜,从而在演化中取得优势。久而久之,合作就会瓦解。
这个困境可以用博弈论中的“囚徒困境”来描述。设想你和一个伙伴面临两个选择:合作或背叛。最好的结局是你背叛而对方合作,你获得所有好处不付代价(得5分);最坏的结局是你合作而对方背叛,你付出一切却什么也得不到(得0分);如果双方都合作,各得3分;如果双方都背叛,各得1分。
从个体理性的角度,无论对方怎么选,你背叛总是更划算:如果对方合作,你背叛得5分胜过合作得3分;如果对方背叛,你背叛得1分胜过合作得0分。所以理性的选择是都背叛,但结果是双方都只得1分,远不如双方都合作各得3分。这就是困境所在。
现实中的很多互利共生关系确实面临这样的威胁。在油菜花田里,有些昆虫会从花朵侧面咬破偷取花蜜,根本不碰花粉,这就是典型的背叛行为。它们得到了食物,但没有为植物提供传粉服务。但为什么这样的背叛没有摧毁整个传粉系统?
降低背叛的吸引力
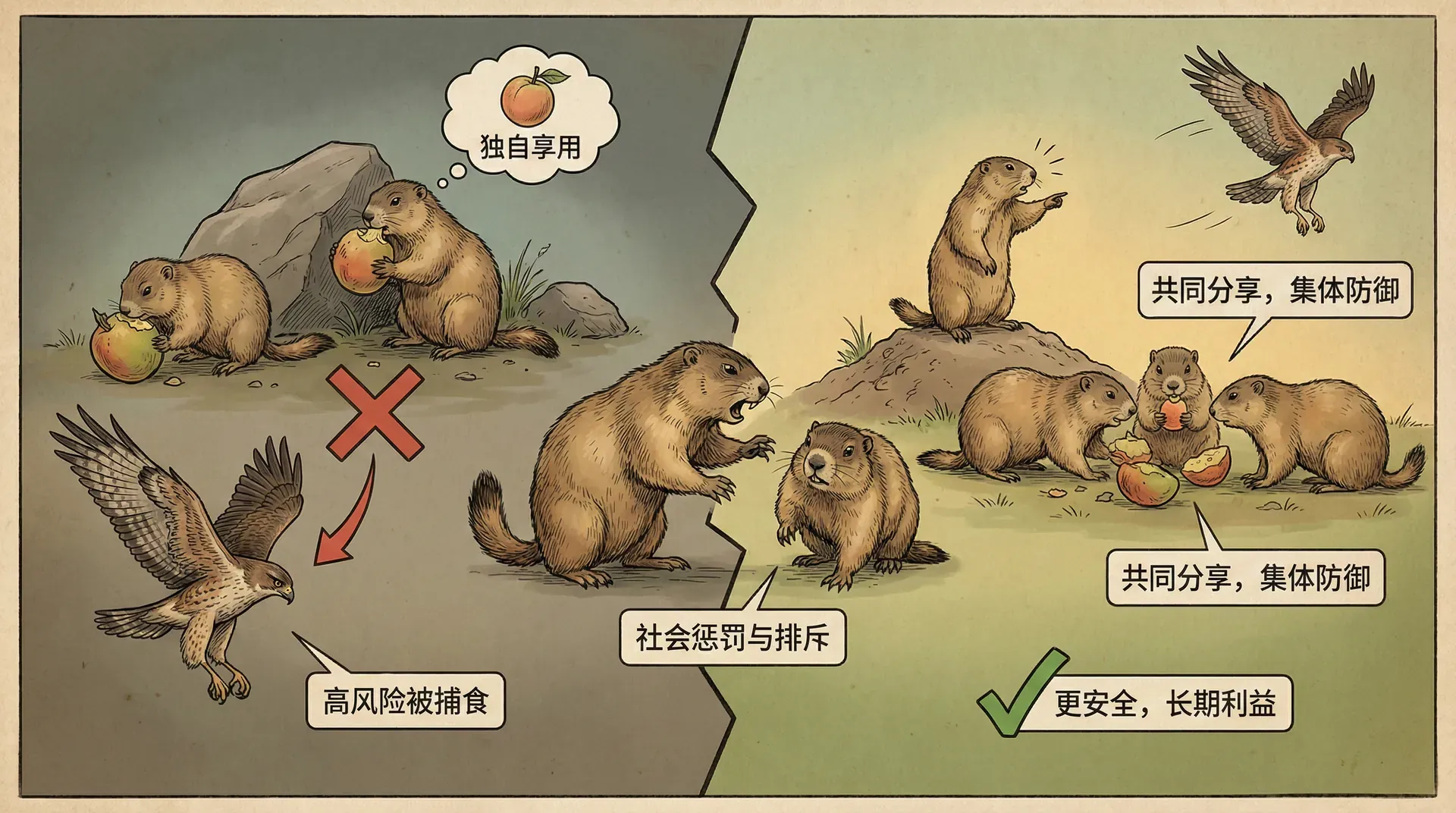
第一个维持合作的机制是:付出的东西本身对自己价值不大。很多互利共生关系中,一方提供的资源对自己来说是过剩的或成本很低的。
植物为传粉者提供的花蜜,主要成分是糖。对于通过光合作用制造糖的植物来说,糖通常不是限制因素,限制植物生长的往往是氮、磷等矿物质。分泌一些糖做花蜜,成本相对较低。同样,植物与菌根真菌的合作中,植物提供碳水化合物(光合作用产物),而获得矿物质和水分,这些正是植物更需要的。
如果提供资源的成本很低,那么即使偶尔遇到背叛者,损失也不大。而只要大部分时候能遇到合作者,整体上还是划算的。这样一来,合作就能维持下去。
制裁与报复
第二个机制是制裁。如果一方能够识别背叛行为并进行惩罚,就能遏制背叛的蔓延。
丝兰蛾不仅为丝兰传粉,还在花中产卵,她的幼虫会吃掉一部分种子。这是一个有趣的平衡:丝兰允许蛾子吃一些种子,作为传粉服务的报酬。但如果某朵花中的幼虫太多,吃掉的种子太多,丝兰就会主动放弃这朵花,让它枯萎脱落。这个举动对丝兰自己也有损失(失去一朵花的所有种子),但它惩罚了过度剥削的蛾子后代,维护了整体的平衡。
豆科植物也有类似的手段。根瘤菌固氮的效率有高有低,有些菌株勤劳,有些偷懒。研究发现,植物能够识别哪些根瘤固氮效率高,哪些低,并相应地调整对这些根瘤的氧气供应。固氮效率低的根瘤得到的氧气少,里面的细菌生长受限。这种「按劳分配」的机制,让偷懒者付出代价,鼓励了勤劳的菌株。
公平分配与隔离
第三个机制是控制繁殖公平性。在有些共生关系中,宿主严格控制共生体的繁殖,确保不同品系的共生体有同等的传递机会。这样一来,任何品系要想成功,唯一的办法就是让宿主成功。
我们自己的细胞就使用这个策略。细胞核里有很多条染色体,每条染色体在细胞分裂时都会被精确复制一次,并且平均分配到两个子细胞中。没有哪条染色体可以通过多复制几次来占便宜。在这种严格控制下,染色体之间没有竞争,唯一的“成功策略”就是让整个细胞成功。这种机制维持了基因组内部的和谐。
一些昆虫和它们的共生细菌也采用类似策略。在某些吸血昆虫体内,共生细菌被分为两组:一组在昆虫的体细胞中工作,帮助消化血液、合成营养物质,但永远不会传给下一代;另一组被隔离在生殖细胞中,负责传递给后代,但不参与日常工作。工作的那一组细菌知道自己不会传给下一代,唯一的“希望”就是让宿主活得好、繁殖得多,因为宿主繁殖多了,那些负责传递的细菌也会繁殖多,而这些细菌和它们有亲缘关系。这种精巧的分工和隔离,有效地遏制了内部竞争。
总结
从稻田里的害虫到根瘤里的细菌,从血吸虫的侵扰到蜜蜂的辛勤劳作,生物之间的关系千姿百态。这些关系不是固定不变的,而是在漫长的演化历程中不断调整、不断塑造的。
理解生物关系的演化,帮助我们认识到:今天看似和谐的共生关系,可能经历了漫长的磨合甚至冲突;今天看似凶恶的寄生,其实也在受到多种因素的制约,可能朝着更温和的方向演化。自然界的智慧在于,它不仅创造了竞争和冲突,也创造了合作和共赢,并且这两者往往交织在一起,构成了生命世界的复杂图景。
当我们下次走进田野,观察身边的动植物时,不妨多想一想:它们之间是什么关系?这种关系是如何形成的?未来又会如何演变?这些问题的答案,就藏在我们周围的每一个生态系统中。