物种是如何诞生的
在地球上,生命的多样性让人惊叹。从喜马拉雅山脉的雪豹到南海珊瑚礁的热带鱼,从东北的红松到西南的望天树,每一个物种都有自己独特的形态和生活方式。这些物种从何而来?它们又是如何产生的?
当我们观察生物的演化时,会发现两个基本过程在起作用。第一个过程叫做渐进演化,指的是一个物种内部随时间发生的改变。比如一种鸟的喙可能在几代之后变得更长或更短,以适应不同的食物。第二个过程是物种形成,也就是一个物种分裂成两个或更多个新物种。正是这两个过程共同作用,才创造出了地球上令人眼花缭乱的生物多样性。
渐进演化创造了物种之间的差异,而物种形成则增加了物种的数量。两者缺一不可,才能解释生命的多样性。
新物种诞生的几种方式
在生命的历史长河中,新谱系(也就是新的物种分支)的产生主要有四种方式。第一种是从无机化学物质组装出生物实体,这个过程目前看来只发生过一次,那就是最初的生命起源。第二种是两个不同谱系通过共生融合成一个新的生物实体,比如线粒体和叶绿体最初都是独立的细菌,后来被纳入真核细胞中。这种情况虽然发生次数不多,但每次都带来了重大影响。
第三种和第四种方式是我们这里要重点讨论的:杂交和谱系分裂。杂交是指来自不同物种的两个个体产生了能够生存和繁殖的后代,并且这些后代与它们的祖先形成了生殖隔离。谱系分裂则是一个物种分裂成两个或更多个物种。
在研究物种形成时,科学家们关注三个方面的变化:生殖隔离(不同物种之间不能杂交)、遗传分化(基因组的差异)和生态分化(生活方式和环境的差异)。这三者有时同步发生,有时一个导致另一个,有时某个变化会显著滞后。
通过杂交产生新物种
植物中的杂交物种
植物界中,通过杂交形成新物种的现象相当普遍。据估计,大约11%的植物物种是通过杂交演化而来的。我们身边就有很多这样的例子。
柚子、橙子和柠檬这些我们熟悉的水果,它们之间存在复杂的杂交关系。橙子实际上是柚子和橘子的杂交种,而柠檬又是橙子和香橼的杂交后代。这些杂交种不仅能够存活,还发展出了独特的特性,比如不同的果皮厚度、酸度和香气。
杂交产生新物种主要有两条途径。第一条途径叫做“重组模型”。在这个过程中,两个亲本物种杂交时,染色体发生断裂和重组,形成全新的染色体排列方式。这些新的排列方式让杂交后代与亲本之间产生了生殖隔离。同时,某些新的染色体组合还可能带来选择优势,让杂交种能够侵入新的生境。
在最初的几代中,杂交种的适应性可能较低,因为基因组合还不稳定。但随着时间推移,有利的基因组合被保留下来,杂交种的适应性可能会超过亲本物种,使它们能够在新的环境中生存。
多倍体化
第二条途径是多倍体化。这是一个特别有意思的过程。正常情况下,每个物种都有固定数量的染色体组。人类有两组(23对),称为二倍体。但有时候,杂交后代可能会获得双倍甚至更多的染色体数量。
我们今天种植的普通小麦是六倍体,也就是说它拥有六组染色体,总共42条。这个六倍体小麦是通过两次杂交和多倍体化事件形成的:首先是两个二倍体野生种杂交形成四倍体,然后这个四倍体又和另一个二倍体种杂交,最终形成了六倍体的普通小麦。
多倍体化为什么能产生新物种呢?关键在于它解决了杂交种的不育问题。通常,来自不同物种的染色体在减数分裂时无法正常配对,导致杂交种不育。但如果染色体数量加倍,每条染色体都能找到自己的配对伙伴,减数分裂就能正常进行,杂交种也就能够繁殖了。
杂交种的适应性之谜
长期以来,科学家们认为杂交种的适应性通常比亲本物种低,因此很难在自然界中长期存在。但近年来的研究改变了这一看法。
在某些环境条件下,杂交种的适应性可能不亚于甚至超过亲本。这种情况在环境发生剧烈变化时尤其明显。当气候或环境突变时,原本适应旧环境的纯种可能面临困境,而具有混合特征的杂交种反而可能因为拥有更广泛的遗传变异而获得生存优势。
环境变化可能为杂交种创造机会。在稳定环境中处于劣势的杂交种,在环境剧变时可能反而成为赢家。
即使在正常情况下适应性较低,一些杂交种群仍然能够长期存在并传播。这说明在特定的生态位中,杂交种可能找到了适合自己的生存空间。
地理隔离与物种分化
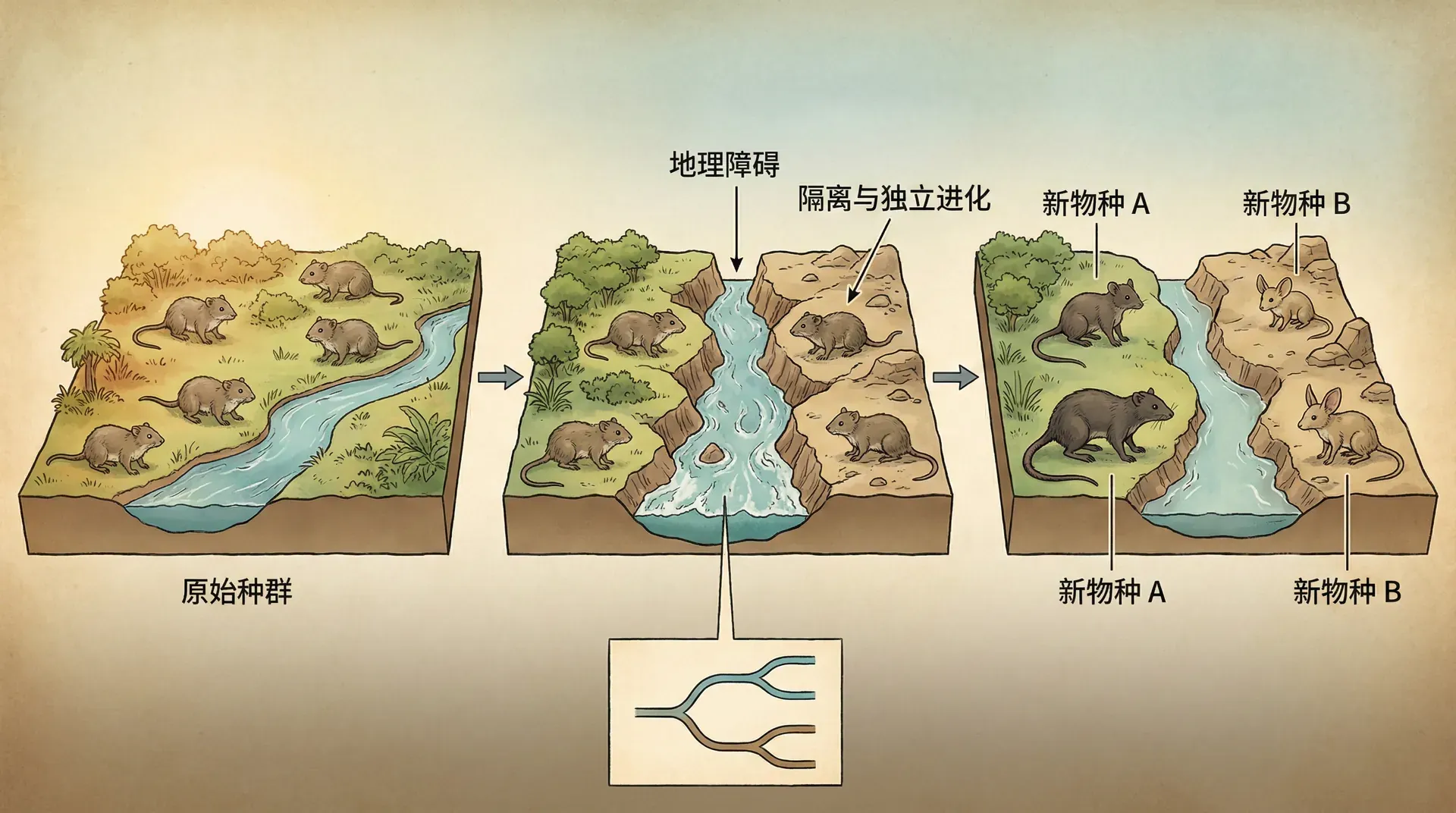
大多数新物种的产生都涉及地理隔离。当一个物种的不同种群被地理障碍分隔开后,它们会沿着各自的路径演化,最终可能演化成不同的物种。这个过程的地理背景可以分为几种不同的模式。
物种形成的地理模式
中国的地理环境为我们提供了观察这些模式的绝佳机会。喜马拉雅山脉和青藏高原的隆起就是一个巨大的地理分割事件。大约从5000万年前开始,印度板块与亚洲板块的碰撞造成了喜马拉雅山脉的抬升。这个过程不仅改变了亚洲的地形,也导致了许多物种的分化。
原本连续分布的物种被这个不断升高的山脉分隔在南北两侧。南侧保持了温暖湿润的气候,而北侧则逐渐变得干旱寒冷。在这两种截然不同的环境中,原本同一物种的南北种群各自适应当地环境,逐渐累积差异,最终形成了不同的物种。今天,我们可以在喜马拉雅山脉南北两侧找到许多这样的姐妹物种,它们形态相似但已经不能杂交。
海洋物种分化
黄海、东海和南海虽然在今天看起来是连通的,但在冰期时,海平面下降可能使得某些海域之间形成障碍。一些鱼类、虾类和贝类的近缘种分布在不同的海域,它们的分化很可能就与这些古老的海平面变化有关。
从图中可以看到,过去200万年间海平面经历了剧烈的波动。在冰期,海平面可以下降100多米,许多浅海区域会暴露出来成为陆地,海洋生物的种群因此被分隔。当间冰期到来,海平面回升,这些种群重新接触时,可能已经演化成了不同的物种。
远距离扩散
间断分化通常涉及远距离的扩散事件。假如一群鸟被风暴吹到远离大陆的岛屿上,或者一些植物种子偶然漂流到新的陆地。这些创始者数量很少,它们携带的基因只是原种群的一小部分。在新环境中,这个小种群会面对不同的选择压力,加上基因漂变的作用,可能会快速演化出新的特征。
南海诸岛就是观察这种模式的理想地点。许多岛屿上的生物显然是从大陆或其他岛屿迁移而来,但经过一段时间的隔离后,它们已经发展出了与大陆种群不同的特征。
同域物种形成
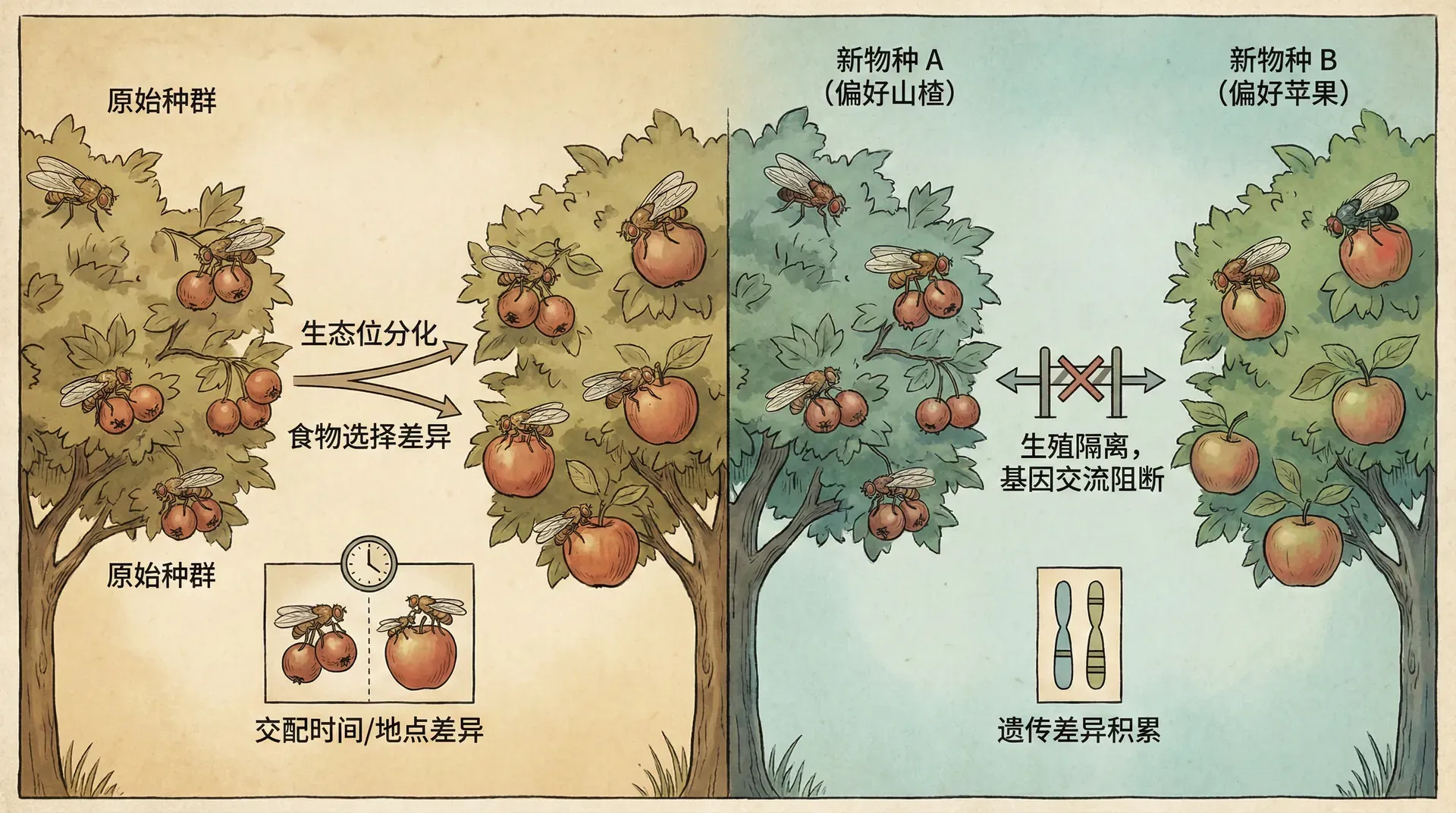
长期以来,科学家们争论一个问题:新物种能否在没有地理隔离的情况下产生?也就是说,在同一个地方,一个物种能否分裂成两个物种?这个问题看似简单,实则困难重重。
理论上的障碍
同域物种形成面临两个主要难题。首先是交配隔离问题。如果两个正在分化的类型生活在同一地点,它们还是会相互交配,基因会持续交流,这会阻止它们进一步分化。其次是生态竞争问题。即使出现了两个生态类型,其中一个往往会在竞争中战胜另一个,而不是两者共存。
但近年来的理论研究发现,如果满足某些条件,同域物种形成是可能的。这些条件包括:强烈的分化选择(偏好两个极端类型而不是中间类型)、交配偏好与生态特征的关联、以及频率依赖的选择(稀有类型具有优势)。
豆象的故事
豆象是一类以豆科植物种子为食的小型甲虫。在中国,绿豆象和赤豆象是两个近缘种,它们分别偏好绿豆和赤豆作为寄主。有意思的是,这两个物种很可能就是通过同域物种形成过程演化而来的。
豆象的关键特征是“寄主忠诚”:雌虫倾向于在自己幼时取食的豆子上产卵。这就创造了一种自然的交配隔离机制。生长在绿豆上的豆象倾向于在绿豆上相遇和交配,而生长在赤豆上的豆象则在赤豆上交配。同时,适应不同寄主的基因也在起作用:适应绿豆的基因在赤豆上可能不那么有效,反之亦然。
图表清楚地显示了两种类型的豆象如何在各自的寄主上逐渐提高适应性,同时在另一种寄主上的适应性下降。这种“权衡”是维持两个生态型分化的关键。杂交个体表现为中间型,在两种寄主上都不如纯种,因此被自然选择淘汰。
性选择的作用
性选择也可能促进同域物种形成。当雌性对雄性的某些特征有强烈偏好时,这种偏好可能会促进种群分化。如果不同雌性偏好不同类型的雄性,而这些偏好又能遗传给后代,那么种群就可能分裂成两个或多个交配系统。
在某些情况下,稀有表型的雄性可能因为竞争较少而获得交配优势。这种“稀有优势” 可以维持多种表型在种群中共存,并可能最终导致生殖隔离的演化。
地理范围重叠的模式
如何判断一个物种是通过同域还是异域方式形成的?一个方法是比较系统发育树上姐妹物种的地理范围重叠程度。如果是同域物种形成,新形成的姐妹种应该有很大的地理范围重叠。如果是异域形成,刚分化时应该几乎没有重叠。
通过这种方法对多个类群的研究显示,异域物种形成确实是主导模式,但同域物种形成在某些类群中也确实存在。
适应性分化的重要性
新物种的遗传分化是如何产生的?是随机的遗传漂变和创始者效应,还是自然选择的作用?这个问题在科学界引发了长期的争论。近年来的证据越来越倾向于后者:自然选择在物种形成中扮演着关键角色。
生态分化与生殖隔离同步
一个有力的证据来自淡水鱼类的研究。在长江和黄河流域,许多湖泊和河流中都生活着鲤科鱼类。在某些湖泊中,我们可以发现两个非常相近的物种共存,一个是底栖型(在湖底觅食),另一个是浮游型(在水体中层捕食浮游生物)。
这两个生态型的分化速度很快,而且生态分化和生殖隔离几乎同时出现。底栖型发展出了下位的口和较粗壮的体型,以便在湖底挖掘食物;浮游型则有上位的口和流线型的体型,便于在水中追捕浮游动物。更重要的是,这两种形态差异提高了各自的觅食效率,这是自然选择的直接结果。
杂交种的适应性检验
如果物种间的差异是适应性的,那么杂交种应该在亲本的自然栖息地中表现不佳,但在实验室中可能表现正常。这是因为实验室消除了自然选择的压力。相反,如果差异是由遗传不兼容引起的,那么即使在实验室中杂交种也应该表现不良。
实际研究发现,许多物种对的杂交种在实验室中是完全可育的。比如云南的几种土著鱼类,它们在自然界中很少杂交,但在实验室条件下却能产生健康的后代。这说明它们之间的隔离主要是生态性的,而非遗传不兼容。
快速的选择响应
对物种间遗传变异的分析也支持自然选择的重要作用。当研究者比较近缘物种或同一物种不同种群之间的基因差异时,发现许多基因位点的变化速度远超中性演化的预期。这说明自然选择在推动这些基因的变化。
有些特征的演化速度快得惊人。在人为改变某些环境条件后,一些物种可以在几代之内就出现显著的适应性变化。这种快速响应只有在强烈的自然选择下才可能发生。
遗传漂变和创始者效应并非不重要,但它们的作用可能被早期研究者高估了。在大多数情况下,自然选择才是驱动物种分化的主要力量。
总结
回顾整个物种形成的过程,生态学的作用贯穿始终。
-
生态环境为物种形成设定了地理舞台。在同域物种形成中,分化选择必须有生态学基础。对于杂交物种形成,地理上的接近是前提,而杂交种的适应性和扩散完全取决于生态条件。即使在异域物种形成中,虽然地理隔离是初始条件,但自然选择仍然经常在创造种群间的初始差异。
-
即使随机过程(如遗传漂变和创始者效应)在某些情况下可能发挥作用,它们也依赖于特定的生态环境,比如小种群规模和地理隔离。
-
自然选择是推动物种间差异不断积累、直至分化不可逆转的主要力量。这个过程本质上是生态学的:不同的环境选择不同的特征,不同的生态位支持不同的物种。
物种的诞生有着深刻的生态学背景。理解物种如何形成,就是理解生物如何与环境互动,如何在演化过程中探索生存的不同可能性。
生命的多样性不是偶然的,而是演化力量在漫长时间里作用的结果。每一个物种的存在,都讲述着一个适应和分化的故事。当我们深入理解这些过程,我们不仅能更好地认识生命的历史,也能更明智地保护今天的生物多样性。