生命演化的证据
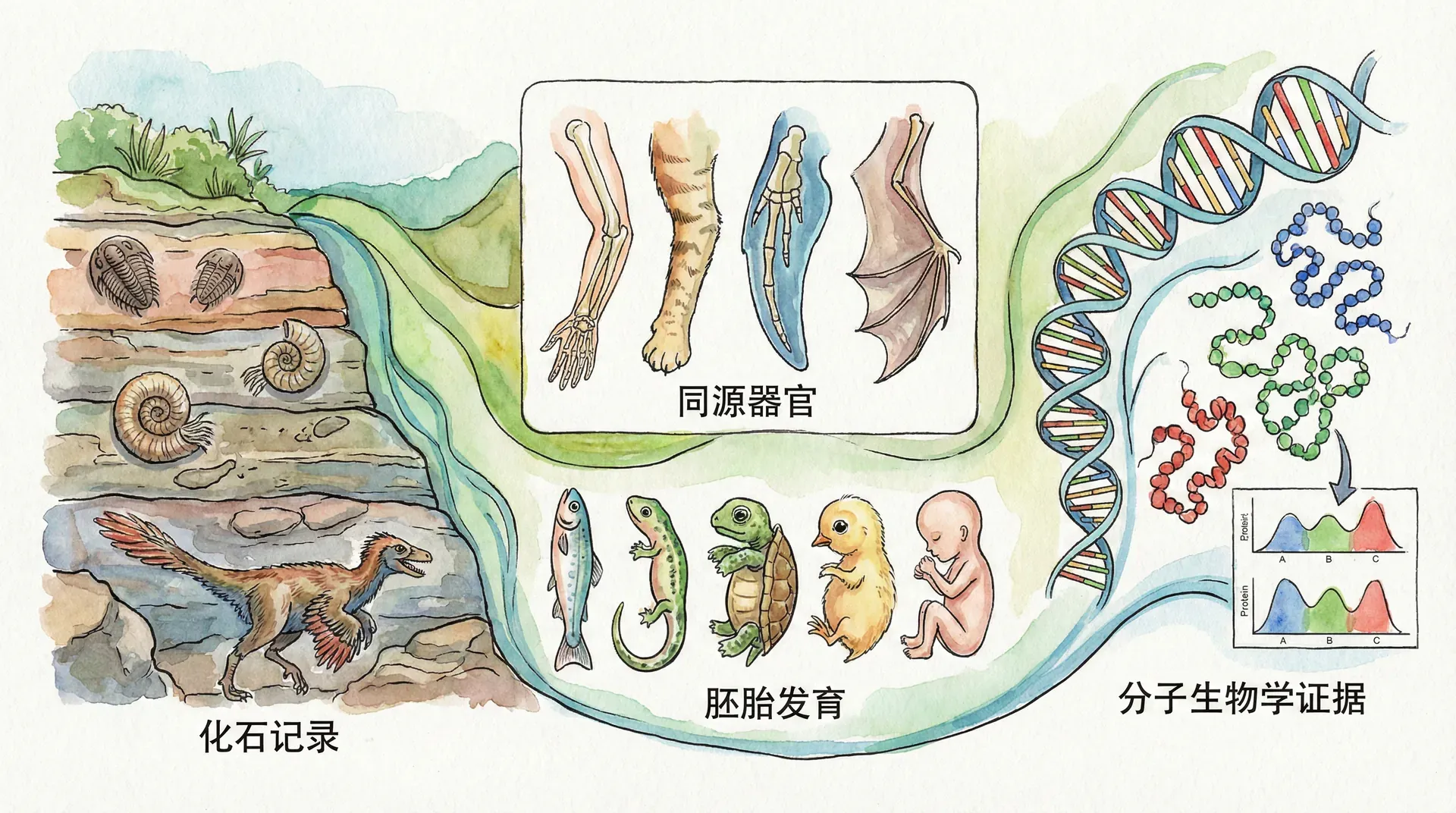
生物进化理论自十九世纪提出以来,历经达尔文、华莱士等科学家的不断完善和现代分子生物学、基因组学的深刻补充,已经成为现代生物学的核心理论框架。它不仅揭示了生物世界的统一性与多样性本源,还为医学、农业、生态保护等诸多领域提供了理论依据。中国幅员辽阔、地质地貌多样,是全球重要的生物演化研究“天然实验室”。本文将结合中国丰富的地质和生物资源,深入考察支持生物进化的多种证据,帮助我们全面理解生命在地球上的演化历程。
生物进化不是一个抽象的、无法验证的理论,而是可以通过大量具体科学证据加以阐明和验证的客观事实。从化石的分布、形态演变,到现存物种的遗传分析和分布格局,再到人工选择带来的剧烈变迁,无一不为进化提供直接或间接的证据。中国在澄江、热河、山旺等地的化石发现,以及对多样生态环境中生物的深入研究,让进化理论的证据显得尤其生动而说服力强。
地质时间与化石记录
地球年龄的认识
理解生物进化,首先要建立对地球历史时间尺度的正确认识。现代科学研究表明,地球大约形成于45亿年前,而最早的生命证据可以追溯至约38亿年前。巨大的时间跨度为生命从简单到复杂的长期演化提供了必要条件。由此,各个地质年代留下了特有的生物遗迹,见证了生命演化的重大事件。
中国的地质研究为地球和生命历史的厘定做出了重要贡献。例如,华北克拉通发现的古老锆石,其年龄超过38亿年,为研究早期地壳形成和生命起源提供了不可替代的证据。此外,中国地质学家主导的北方盾构及青藏高原抬升等重大事件的年代厘定,也为地球历史重建提供了基石。
为了直观了解地质年代及代表性生物事件,下面通过图表进行总结:
化石记录的不完整性与独特性
化石的保存需要特殊环境(如突然埋藏、缺氧环境等),因此全球范围内的化石记录本身就是极不完整的,但这并不妨碍科学家通过现有化石还原生物演化的宏观图景。中国在古生物研究领域具有独特优势,不仅发现了大量数量和类型丰富的化石,而且保存状况极佳,有些甚至见证了生物“群爆发”与“大灭绝”等现象。
例如,云南澄江生物群出土了距今约5.3亿年前寒武纪早期的精美化石,该地化石成功保存了大量软体生物和节肢动物,部分个体甚至连细胞结构、消化道等软组织都得以完整展现,极大丰富了我们对生命早期演化的认知。和澄江一样,山东山旺中新世植物化石丰富,辽西热河生物群保留了恐龙和鸟类羽毛等精细结构,贵州关岭三叠纪产地则完整展现了海生爬行动物的生态。
这些研究不仅让我们得以窥见远古生命原貌,还让科学家能分析不同地层的生物群落结构变化,追踪生物门类的起源、扩散乃至多次大灭绝与复苏的进程。
化石记录中的演化模式
尽管化石记录永远不会“无缝衔接”,但通过各时期化石比较,依然能够识别出生物进化的清晰轨迹。以中国境内发现的脊椎动物化石为例,从寒武纪最早的原始鱼类、志留纪的有颌鱼、泥盆纪的四足动物,到后来两栖类、爬行类、恐龙、鸟类及哺乳动物,脊椎动物展现了由简单向复杂、由水生向陆生、由单一向多样的演化趋势。
不仅如此,无脊椎动物同样经历了多样化和复杂化过程。两者间复杂度指数随时间演进的对比,可以通过下方图表直观体现:
通过分析这些复杂度指数的变化,可以看到新型生物结构和功能不断涌现,不同生物门类相继崛起,响应着环境变迁、气候波动及大陆漂移等地球历史事件。这些都是生物进化在地质时间尺度上客观发生的直接证据,也让人类得以推断自己与其他生命共享的演化历史。
共同祖先的证据
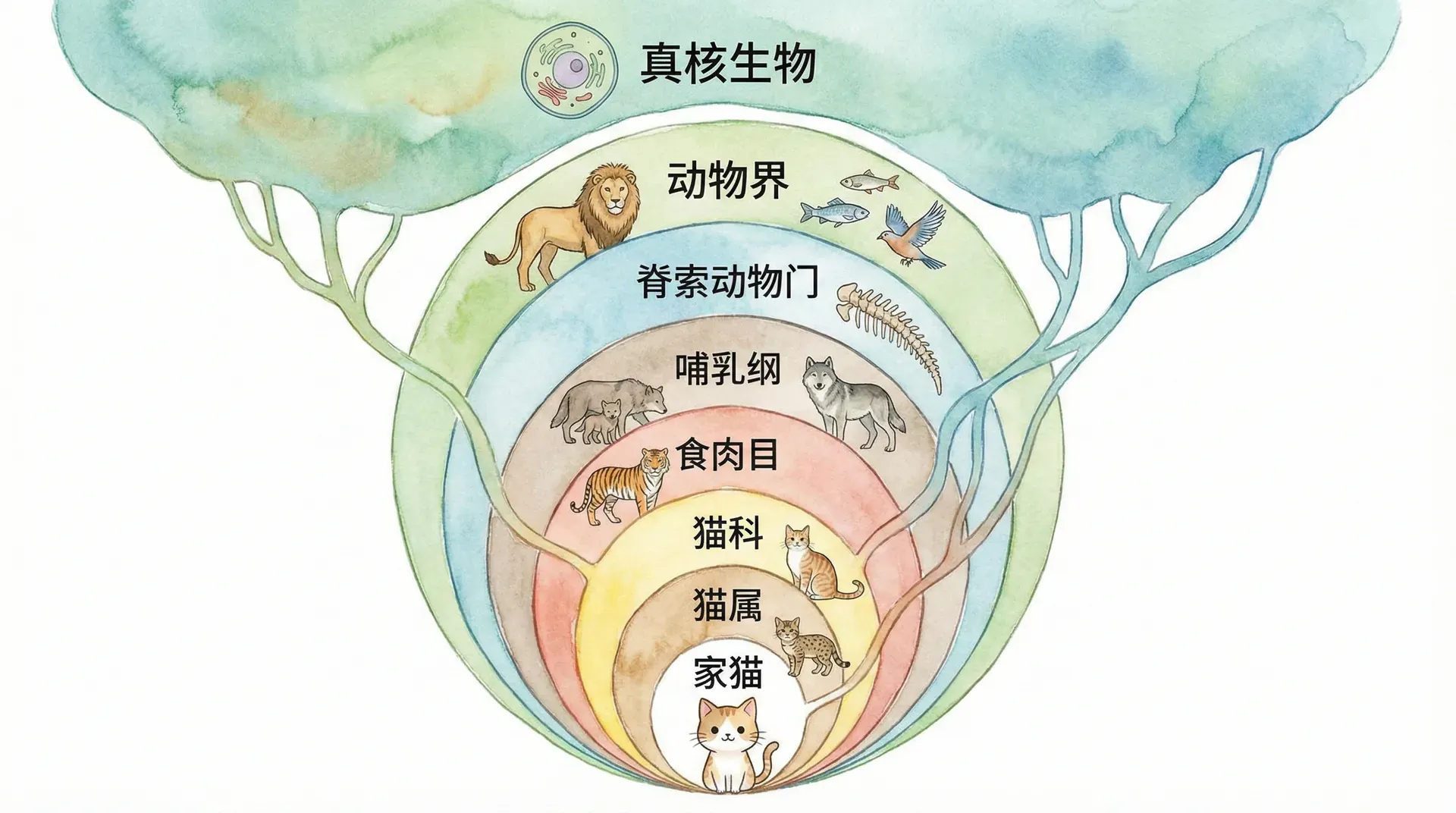
分类系统的层次结构
所有生物都可以按照相似性、形态特征和遗传信息分成不同的等级群体,形成一个彼此嵌套、包含关系明确的系统层次结构。这种方式最初由18世纪瑞典博物学家林奈建立(称为“林奈系统”),最初依赖外部形态,后来随着进化论和分子生物学的发展,这一体系得到了大量证据的支持与完善。层次结构不只是分类的便利,更反映了物种间真实的亲缘关系。
这种嵌套关系的本质在于:如果所有生物都起源于共同祖先,那么随着时间推移,祖先种群不断发生分化与演化,每个分支再继续分化,最终形成现代的多样生命,正所谓“一树生万枝”,各类群之间的包容和分化时间可通过分类体系明确标注。
让我们以中国的猫科动物为例来理解这种层次关系,并补充一些常见的分类情况:
在中国境内生活着多种猫科动物,包括东北虎、华南虎、雪豹、猞猁、豹猫、金猫、兔狲等。虽然它们的体型、花纹、栖息环境各不相同,但都具有猫科动物的基本特征:尖锐的犬齿、可伸缩的爪子、优秀的夜视能力等。此外,无论大型的东北虎还是小型的豹猫,它们都保留了祖先的许多核心遗传信息,以及骨骼、牙齿、听觉等方面的同源性。
通过不同动物的分层级亲缘关系,我们可以直观理解共同祖先理论:每一层(纲、目、科、属、种、亚种)都是一次演化“分家”。今天中国的猫科动物正是祖先种群不断演化、适应不同生态位积累的结果。类似的树状层次,也普遍适用于所有生物门类,不仅限于哺乳动物。
分子证据支持共同祖先
随着分子生物学和基因测序技术的发展,我们获得了比形态更直接的物种关系证据。通过对不同物种的DNA、RNA和蛋白质序列进行比较分析,可以建立“分子系统树”或“系统发育树”,其结构与传统形态分类高度吻合,也为共同祖先理论提供了强有力支持。
基因相似性和单一祖先理论
以熊科动物为例,大熊猫、亚洲黑熊、棕熊、北极熊等虽然外观差别极大,但通过线粒体DNA和细胞核基因组的测序发现,它们无一例外都保留着高度同源的关键基因(如线粒体细胞色素b基因等)。相同或相似的突变累积顺序,直接反映了不同分支的分化时间和亲缘距离。
基于类似的遗传数据,不仅熊科、猫科等哺乳动物展现出清晰可辨的分支关系,甚至中国和全世界的鸟类、两栖类和鱼类也能通过分子钟方法推算分化年代。研究发现,分子数据与地质化石资料、形态学证据高度吻合,支持了生物的普遍共同祖先理论。
不仅如此,一些分子“遗迹”也支持共同祖先模型。例如,人类和其它哺乳动物基因组中均存在若干“无用基因”(伪基因),这些基因已经失活,但在系统进化树中具有同源起源,说明它们都来源于共同祖先基因组中的原始基因。
地理分布的规律
岛屿生物地理学
地球表面并非均质的生物舞台。空间隔离与地理屏障促使同类祖先分化为不同物种,形成“地理种化”现象。中国拥有众多岛屿和群岛,为我们研究生物地理分布和演化机制提供了天然实验场。
以海南岛、台湾岛等大型岛屿为例,其动植物组成明显区别于大陆和彼此之间,反映了岛屿独特的地质历史与隔离进程:
海南岛的哺乳动物群落与大陆华南地区高度相似,这是因为在冰期海平面下降时,海南岛曾与大陆完全连通,许多动物在这一时期迁徙定居。反观台湾岛,由于比海南岛更早与大陆分离,隔离时间更长,岛上特有种比例更高,包括台湾黑熊、金翼白眉等,反映了较多独立进化事件。
此外,崇明岛、舟山群岛等新生岛屿由于隔离时间较短,特有种数量和比例都显著偏低,说明物种分化需要较长的地理隔离时间。
岛屿的生物多样性差异印证了演化论中的“地理隔离种化”机制。随着孤立时间拉长,岛屿生物在不同环境压力下渐渐形成独特的新物种,这一规律在中国众多岛屿和湖泊中屡见不鲜。
大陆漂移的证据
除了岛屿地理隔离,地壳大规模运动(板块漂移)在地球历史长河中也极大影响了生物分布。中国丰富的地质遗迹和化石数据,反映了一系列由于板块漂移造成的生物区系变化。
例如,侏罗纪时期中国与其他大陆板块相对独立,出现了许多独特的“东方生物区系”。进入新生代,板块聚合和抬升促成青藏高原的形成、大型湖泊与山脉孤立,为动植物的快速分化和区系更替提供了机会。
由上述图表可见,每个时期新形成的区系数量与累积总数都与大陆板块运动密切相关。例如青藏高原的隆升创造了横断山脉这样多样化的生态走廊,大幅度增加了中国境内的生物多样性。曾经的“联通”或“分裂”,不仅可以通过地质证据得到印证,也通过转移物种化石分布和现代区系特征得以事实验证。这些都证实大陆漂移和地球历史事件是驱动生物进化格局的重要因素。
比较解剖学的证据
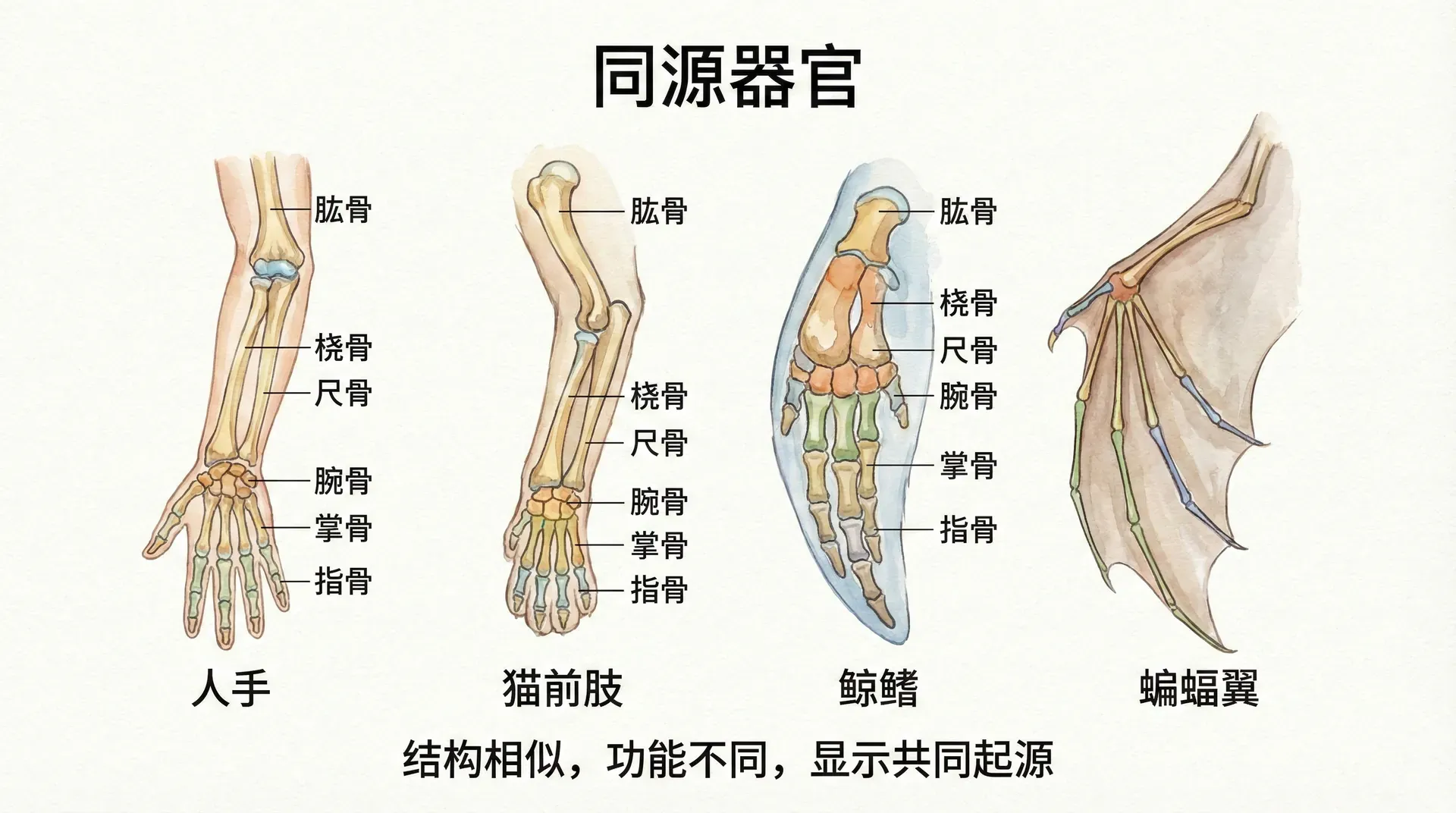
同源器官的相似性
如果不同生物确实拥有共同祖先,它们身体结构的深层共性就不足为奇。所谓“同源器官”,指的是由同一祖先生物的同一结构演变来,但在不同物种间功能可能发生巨大变化的结构。比较解剖学正是研究这些现象的主要分支。
中国哺乳动物为我们提供了诸多生动典例——即便功能差异极大,不同种类动物前肢的骨骼组成仍然高度一致:
大熊猫的前掌、华南虎的前爪、中华白海豚的胸鳍,虽然功能完全不同(抓握竹子、捕猎、游泳),但它们的骨骼结构却惊人地相似,都包含相同的骨骼元素:肱骨、桡骨、尺骨、腕骨等。这种结构一致性超越了环境和生理需求的差异,是血缘关系的直接证据。
以上同源骨骼,均源自早期哺乳动物前肢骨架,通过自然选择和功能改造,分别适应了陆地、森林、洞穴、河流甚至海洋等不同生态环境。对比鸟类翅膀、鲸鱼鳍肢、蝙蝠翼等同源结构,会发现更广泛、更深刻的进化模式。
痕迹器官的存在
痕迹器官(vestigial organs)是指在部分物种体内残留,却已失去原有功能或功能极大削弱的器官。这些“功能残影”只有通过进化论才合理解释——它们是祖先进化阶段的“遗产”,在环境与适应改变后逐渐退化。
中国动物界有大量可观测的痕迹器官实例:
- 中华白海豚后肢骨:现存白海豚体内埋藏着一对非常小的骨块,实际上是祖先从陆生向水生演化过程中退化的后肢遗迹。虽然已不能用于行走,却依然能被解剖发现。
- 穿山甲牙齿:中华穿山甲是中国特有哺乳动物,其成年个体已无功能性牙齿,口腔内只留下牙床等遗痕。早期穿山甲祖先有完整的牙齿,化石证据表明这一特征经历了数千万年渐进退化。
此外,许多动物体内还保存有胚胎时期才出现,但最终消失或缩小的痕迹结构。例如部分类人猿和人类存在“尾骨”、部分哺乳动物体内隐藏“第三眼睑”等等。这些事实共同说明,不同生物的共同祖先随着环境与生活方式变化,其功能性器官有的强化演化,有的则逐步退化、残留成“痕迹”。
胚胎发育的证据
现代生物学认为,胚胎发育过程中出现的结构和过程的相似性,是揭示不同动物存在共同祖先的重要线索。尽管成体形态、结构差异极大,但在胚胎早期,这些差异往往被高度统一的发育进程所掩盖。
胚胎相似性
不同物种在胚胎发育早期阶段表现出惊人的相似性,这种现象在脊椎动物中尤为明显。例如,无论是鱼类(如中华鲟)、爬行动物(如扬子鳄)、还是哺乳动物(如大熊猫与人),早期胚胎都经历受精卵分裂、囊胚、原肠胚的基本阶段,出现脊索、体节、鳃裂等结构。这些发育阶段的共性证据,为所有脊椎动物来自共同祖先这一观点提供有力支持。
中国科学家对本土代表性物种开展了大量胚胎学研究。例如,利用中华鲟、扬子鳄和大熊猫的胚胎发育过程,揭示了猫、鳄、鱼等多个类群在早期胚胎形态、发育基因激活顺序等方面的相似性。成体阶段差异巨大的动物,竟在最初发育时“难分你我”,这是进化论最令人信服的现象之一。
需要注意的是,现代胚胎学否定了早期极端的“重演律”(即胚胎发育会完整重演进化史),但强调胚胎发育早期出现的相似性,反映了发育调控网络高度保守,以及某些祖先阶段特征的暂时性保留。这为我们认识胚胎的演化保守性提供了更精准视角。
真实的胚胎比较可以在各阶段反映相似与差异。让我们通过数据来观察中国几种脊椎动物胚胎发育的相似指数变化:
可以看出,越早期阶段的相似性越高,随着发育深入,各物种的独特性逐渐显现。这印证了进化过程中,祖先特征在胚胎早期被广泛保留,而特化性状则在后期才表达。中国学者通过分子、生理和形态的多层次分析,为动物进化树的分支关系提供了坚实证据。
发育基因的保守性
近年来,分子胚胎学揭示出,许多调控胚胎发育的关键基因在极为遥远的物种间都高度相似。这些基因家族(如Hox、Pax、Hedgehog、Wnt等)不仅控制了不同生命阶段的重要发育事件,其序列及表达模式保守至今。
例如,在对大熊猫、小鼠等胚胎实施基因敲除实验时,研究发现Hox基因家族的缺陷会导致脊柱、四肢等结构发育异常或畸形;在中华鲟和扬子鳄研究中也证实了Wnt和Hedgehog信号通路对胚胎早期体轴建立和器官形成至关重要。这些现象不仅揭示了发育基因的功能重要性,也佐证了这些基因自远古动物演化而来,至今在物种间仍保持高度一致的进化事实。
分子演化的现代证据
随着DNA测序和生物信息学的发展,分子证据已成为现代进化生物学的核心之一。分子水平的比较为物种间的亲缘关系提供了客观、量化的数据基础。
DNA序列比较
DNA是记录生物遗传信息的分子,通过对比不同物种的DNA序列,科学家可以推断它们之间的进化距离及分支时间。近年来,中国科学家积极参与大熊猫、金丝猴、华南虎等濒危物种的基因组测序项目,极大丰富了我国及全球物种DNA数据。
例如,下图展示了黑猩猩、猕猴、大熊猫、华南虎、中华白海豚、小鼠与人类的基因组相似性:
可见,物种间基因组的相似性与它们的亲缘关系高度吻合。例如,我们与黑猩猩的基因组相似度高达98.8%,但与大熊猫、老虎等相对较远。中国还对本土特有动物的适应性基因、进化相关新基因开展了深入研究,如揭示大熊猫适应低营养竹叶饮食的消化酶基因、华南虎相关适应性的基因变异等。
分子钟的应用
分子钟理论提出,遗传物质的变异在时间上以大致恒定的速率积累,因此通过DNA或蛋白质序列的差异,可以推算物种分化的年代。中国科学家将分子钟技术应用于本土野生动物研究,取得了许多突破性成果。
例如,通过对大熊猫及其近缘熊科物种基因组的分子钟分析,科学家估算大熊猫大约在1900万年前就已经从其他熊科动物中分化出来,这一结论与中国化石记录高度一致,有力佐证了化石与分子两种证据的互补和一致性。
分子钟不仅用于物种起源时间推算,也广泛应用于疾病进化、病毒溯源、家畜家禽驯化等领域,极大丰富了我们对生物历史的认识。
人工选择的实验证据
中国的农业驯化
除自然选择外,人工选择也是驱动短期内物种快速变化的重要机制。中国作为世界农业的发源地之一,为人工选择的效应提供了众多鲜活案例,无论是栽培作物还是驯化动物,演变速度远超野外自然变异。
如水稻通过长期人工选择,籽粒变得更大、不易脱落,极大提高了产量和收获率。家猪和家鸡的体型、性格、产蛋量等都发生了巨大改变。家蚕驯化过程中,野生生存能力丧失,但丝质产量和质量大幅提升。中国科学家通过考古、遗传和分子证据结合,精确还原了这些物种驯化和性状演变的历史轨迹。
除了作物和家畜,人工选择还涵盖观赏鱼、狗、猫、鸽等经济与伴侣动物,以及茶树、苹果、柑橘等园艺作物。中国丰厚的农业传统和多元化的地理环境,使得人工选择和快速演化的实例极为丰富。分析这些变化的遗传基础,进一步证明了选择压力(无论是自然还是人为)都能够导致物种在较短时间尺度上的明显演化。
人工选择所导致的表型差异,有时甚至超过了物种间通过自然选择缓慢积累的变化,这也为现代生物技术(如分子育种、定向改良)提供了理论指导和现实案例。
总结
通过对各类证据的综合分析,我们可以清楚地看到生物进化是一个客观存在的自然过程。从中国丰富的化石记录,到现生物种的地理分布模式,从比较解剖学的发现,到现代分子生物学的成果,所有证据都指向同一个结论:地球上的所有生物都有着共同的祖先,并通过漫长的演化过程形成了今天我们看到的生物多样性。
理解生物进化不仅有助于我们认识生命的本质,更为生物多样性保护、疾病防治、农业育种等实践活动提供了科学指导。中国作为生物多样性最丰富的国家之一,在进化研究方面具有得天独厚的优势。
生物进化理论的确立,标志着人类对生命认识的一次重大飞跃。它不仅改变了我们对生物世界的理解,也深刻影响了我们对人类自身地位的认识。在新的历史时期,我们要运用进化的思维,更好地保护和利用生物资源,实现人与自然的和谐发展。