神经细胞如何传递信息
如果把一个现代人丢到深山老林里,他能活多久?可能从未狩猎过野兽,偶尔钓过鱼就算幸运了。也许从未种植过足够的粮食蔬菜来填饱肚子,更别说搭建遮风挡雨的房屋或缝制御寒的衣物。除了本能的呼吸,还有什么生存技能能够完全独立掌握呢?人类集体协作能创造奇迹,但孤军作战时往往束手无策。
我们神经系统中的细胞也是如此。它们协同工作能完成令人惊叹的任务,但单个细胞却显得渺小无助。在探索神经系统如何协同工作之前,首先要理解单个神经细胞是如何运作的。这就像是理解每个汉字的构造,才能读懂一篇文章的含义。
这部分内容涉及较多生物化学原理,如果对化学基础概念还不太熟悉,建议先回顾一下相关知识,这样能更好地理解神经细胞的运作机制。
神经科学的开拓者
19世纪末之前,人们对大脑的微观结构知之甚少。研究者们甚至无法确定神经纤维究竟是连成一体的网络,还是由彼此分离的独立细胞构成。这个问题悬而未决了很长时间,直到西班牙科学家圣地亚哥·拉蒙-卡哈尔凭借一套巧妙的染色方法,才将答案呈现在世人面前。
拉蒙-卡哈尔原本想成为艺术家,但在父亲坚持下学习了医学。他将艺术天赋与科学研究完美结合,成为了杰出的解剖学研究者和插画家,所绘制的神经系统图谱至今仍被奉为经典。
拉蒙-卡哈尔曾因为上拉丁语课不专心而被关进小黑屋,每天只给一顿饭,还要接受公开鞭打——那时他只有10岁。这位日后的神经科学奠基人,少年时代的经历并不顺遂,却丝毫没有磨灭他对知识的热情。
卡哈尔使用意大利科学家高尔基发明的银染色法,但他巧妙地将其应用于幼体大脑——因为幼体的细胞更小,更容易在显微镜下观察清楚。通过这种方法,他最终证明了神经细胞彼此分离,并非融合成一个整体。这一发现奠定了现代神经科学的基础。
“有多少有趣的事实因为最初的观察者认为它们理所当然而未能转化为富有成效的发现!奇怪的是,那些用巫师传说、神秘事件和非凡现象滋养想象力的民众,却轻视周围的世界,认为它们平淡、单调、缺乏诗意,没有意识到其实一切都充满了秘密、神奇和惊喜。”——拉蒙-卡哈尔
细胞的基本结构
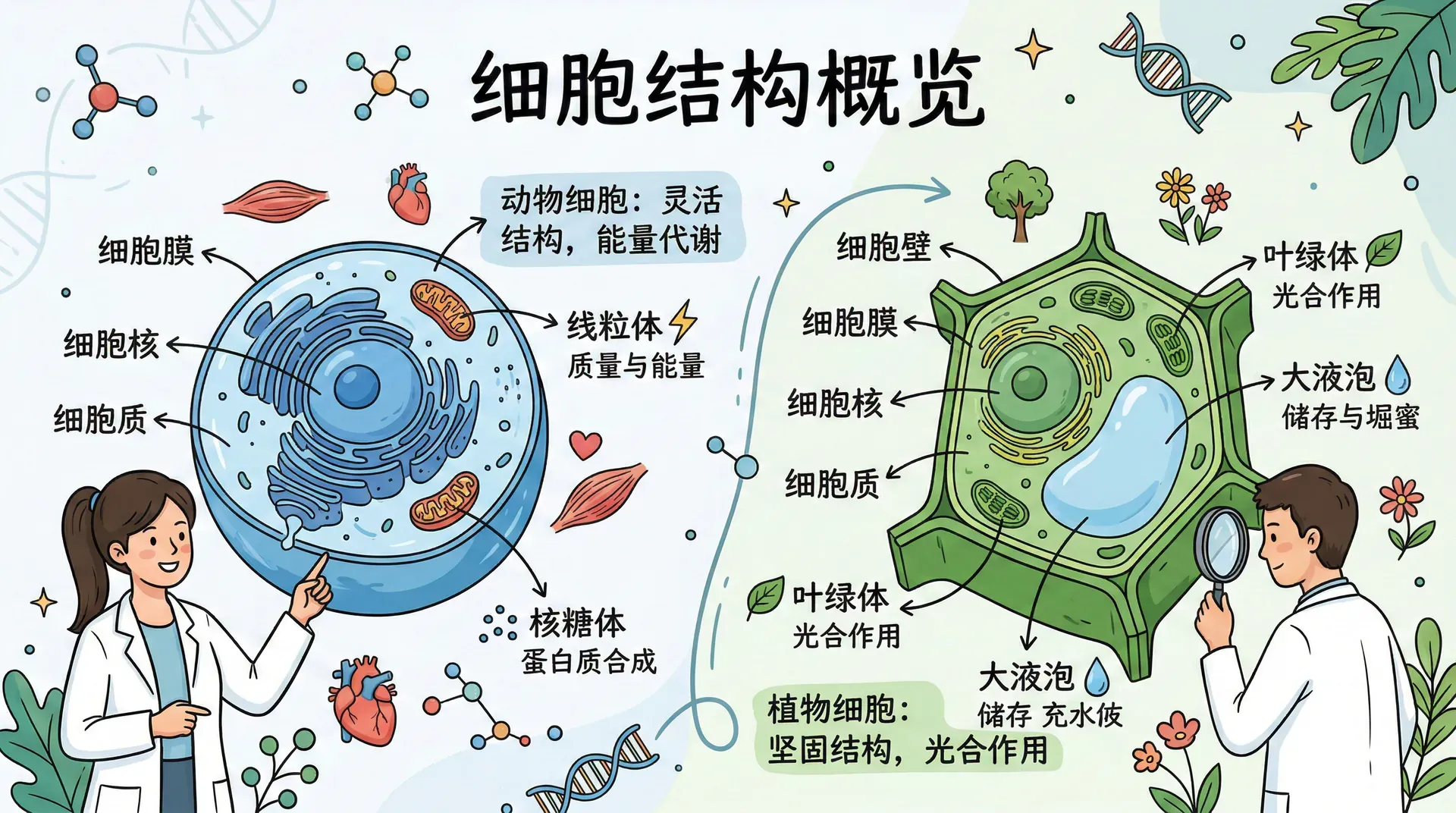
要理解神经细胞的运作,首先需要了解动物细胞的共同特征。神经元虽然在形态上极为特殊,但在细胞生物学的基础层面上,它与其他细胞共享着一套相同的核心结构。
细胞膜是细胞表面的一层约8纳米厚的屏障,厚度还不到头发丝的万分之一。这层膜由两层磷脂分子构成,其中嵌入着蛋白质通道,精确控制各种化学物质的进出。这种双层结构既保证了细胞的相对封闭性,又允许水分、氧气、各种离子等重要物质进行必要的交换。细胞核是存放遗传信息的核心区域,除了哺乳动物的红血细胞之外,所有动物细胞都有细胞核,染色体储存在这里,承载着细胞运作的全部指令。线粒体则负责代谢活动,通过消耗燃料和氧气为细胞的所有活动提供能量。核糖体是合成蛋白质的场所,有些核糖体在细胞中自由漂浮,有些则附着在内质网上,由内质网负责运输新合成的蛋白质到各处。
神经元的结构与类型
神经元最显著的特征是它们千变万化的形状。与大多数其他细胞不同,神经元长出了长长的分支状延伸,这些延伸结构使得神经元可以与远距离的目标建立联系,形成复杂的信息传递网络。
神经元的四个主要组成部分
大型神经元通常由树突、胞体、轴突和突触前末梢四个部分构成,每个部分承担着不同的功能职责。
树突是从胞体向外延伸的分支状纤维,末端越来越细。树突表面布满了专门的突触受体,用来接收其他神经元传来的信号。树突的表面积越大,能接收的信息就越多。许多树突还长有细小的突起,称为树突棘,进一步增加了可用于突触连接的表面积。胞体包含细胞核、核糖体和线粒体,是神经元进行新陈代谢的主要场所。哺乳动物神经元的胞体直径从0.005毫米到0.1毫米不等,而某些无脊椎动物的神经元胞体甚至可以达到整整一毫米。
轴突是一根细长的纤维,直径基本恒定,通常比树突更长。它是神经元向外传递信号的通道,将电信号送往其他神经元、器官或肌肉。许多脊椎动物的轴突外面包裹着绝缘材料——髓鞘,中间有节段性的间隔叫做郎飞结。轴突有许多分支,每根分支的末端都会膨大,形成突触前末梢,这是轴突释放化学物质的关键位置。
一个神经元可以有很多树突,但只能有一个轴突。不过,轴突可以在远离胞体的地方分出许多分支。有些轴突长达一米多,从脊髓一直延伸到脚部,这意味着一个细胞可以跨越相当大的距离来传递信号。
感觉神经元与运动神经元
神经系统中存在两种功能各异的主要神经元类型,它们的结构直接反映了各自承担的工作。运动神经元的胞体位于脊髓中,通过树突接收来自其他神经元的刺激,然后沿轴突将信号传导至肌肉,负责执行大脑发出的运动指令。感觉神经元的一端特化为高度敏感的感受器,能够感知光线、声音或触觉等特定刺激。信号从感受器进入轴突,而细胞体则位于主干旁边的小分支上。
在神经科学中,还有三个常见的方向性术语需要区分:传入轴突将信息带入某个结构;传出轴突将信息带出某个结构;中间神经元的树突和轴突则完全位于同一结构内部。值得注意的是,同一个神经元可能在整体系统中既充当某一结构的传出成分,又充当另一结构的传入成分,这取决于所考察的层级。
神经元形态的多样性
神经元的大小、形状和功能存在巨大差异,这种多样性与各自承担的信息处理任务密切相关。小脑的浦肯野细胞拥有极为广泛的树突分支,能够接收多达20万个输入信号,参与精细的运动协调。相比之下,视网膜中的某些细胞只有很短的树突分支,只能整合来自少数几个来源的信号,专注于处理特定的视觉信息。
连接范围更广的神经元能与更多目标建立联系,而连接较为集中的神经元则在特定的信息处理环节中发挥专门作用。这种多样性共同构成了神经系统处理复杂信息的基础。
胶质细胞的功能
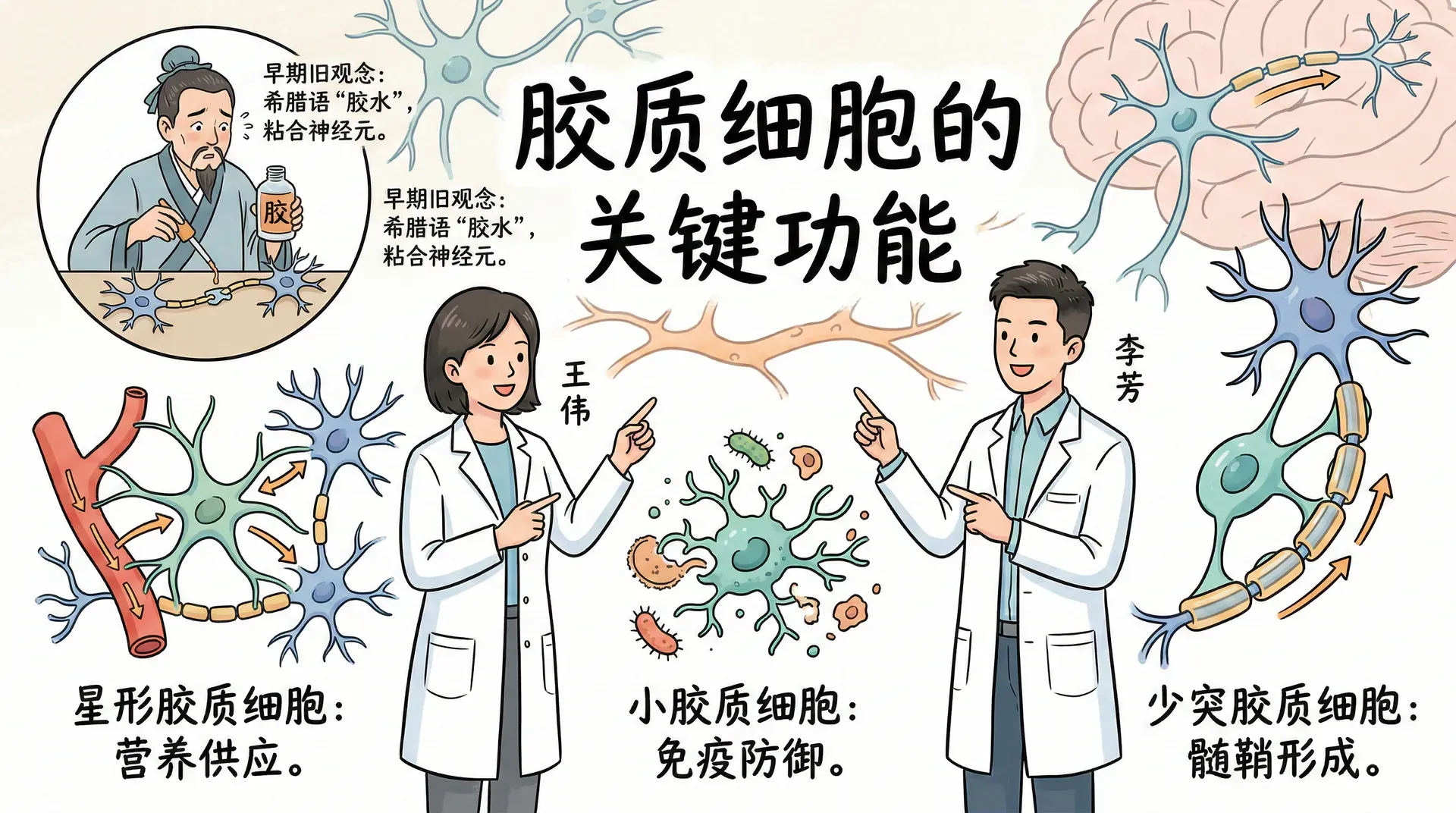
除了神经元,神经系统还包含另一类重要细胞——胶质细胞。它们的名字来源于希腊语中的“胶水”一词,因为早期研究者认为它们将神经元粘合在一起。虽然这种理解已经过时,但名称保留了下来。胶质细胞比神经元小,数量却更多,虽然不负责远距离信号传递,却执行着许多不可缺少的功能。
星形胶质细胞
星形胶质细胞是一类星状的细胞,包围着一群功能相关轴突的突触前末梢。它们通过吸收轴突释放的离子并将其重新释放给轴突,帮助同步这些轴突的活动,使它们能够协调地发送信息。
除了协调功能,星形胶质细胞还承担其他职责。它们负责清理神经元死亡后产生的废物,控制每个脑区的血流量,在某些区域活动增强时扩张血管以输送更多营养物质。它们甚至参与神经递质的回收利用——将神经元释放的谷氨酸转化为谷氨酰胺再传递回神经元,形成一套精密的循环系统。
小胶质细胞
小胶质细胞体积非常小,是大脑的免疫执行者,负责清除废物、病毒、真菌和其他微生物。当神经组织受损或受到外来威胁时,小胶质细胞会迅速聚集到受损区域,清理残留物质并防止炎症扩散。从某种意义上说,它们是免疫系统在大脑内部的驻守代表。
少突胶质细胞与施万细胞
在大脑和脊髓中,少突胶质细胞负责构建包围某些脊椎动物轴突的髓鞘;在身体其他部位,这个任务由施万细胞承担。髓鞘对于轴突信号的快速传导至关重要,下文将详细说明其机制。
一个少突胶质细胞可以为30到50个不同的轴突形成髓鞘段,以高效的方式服务于多个目标,大大节省了神经系统的资源。
放射状胶质细胞
在胚胎发育期间,放射状胶质细胞引导神经元及其轴突和树突的迁移,相当于在发育中的大脑里搭建了一套导向系统。发育完成后,大多数放射状胶质细胞分化成神经元,少数则转变为星形胶质细胞和少突胶质细胞。
血脑屏障
大脑和其他器官一样需要从血液中获取营养,但许多化学物质却无法从血液进入大脑。这种将大多数外来物质拒之门外的机制叫做血脑屏障,是保护神经系统的重要防线。
血脑屏障存在的原因
当病毒入侵细胞时,细胞会将病毒颗粒推出到膜表面,让免疫系统发现并消灭它们。这种策略对皮肤细胞或血细胞来说没有问题,因为身体可以轻易替换受损的细胞。但脊椎动物的大脑很少替换受损的神经元。为了最大限度地降低不可修复的脑损伤风险,身体沿着大脑血管壁建起了一道屏障,阻挡大多数病毒、细菌和有害化学物质进入神经组织。
某些病毒,如狂犬病病毒,能够绕过血脑屏障感染大脑,后果往往是致命的。一旦病毒进入神经系统,就很难被彻底清除。引起水痘和带状疱疹的病毒会潜伏在脊髓细胞中,即使免疫系统在神经系统外有效控制了这些病毒,病毒颗粒仍会留在脊髓中,几十年后可能重新激活。
物质穿越血脑屏障的方式
血脑屏障依靠形成毛细血管壁的内皮细胞发挥作用。在大脑外,这些细胞之间有小间隙,但在大脑中,它们紧密连接,几乎任何物质都难以通过。
这种强大的防御也有其代价:血脑屏障不仅阻挡有害物质,也阻挡部分有用的化学物质,包括某些燃料和氨基酸。为了让大脑正常运作,必须有特殊机制让这些化学物质穿越屏障。
-
小的不带电分子,包括氧气和二氧化碳,可以自由穿越。水分通过内皮细胞壁上的特殊蛋白质通道穿越。
-
能溶解在膜脂质中的分子可以被动穿越,包括维生素A和维生素D,以及多种影响大脑的药物——从抗抑郁药、精神科药物到某些非法药物均属此类。
-
主动转运是一种耗费能量的蛋白质介导过程,将化学物质从血液泵入大脑。通过这种方式进入大脑的物质包括葡萄糖、氨基酸、嘌呤、胆碱、部分维生素、铁和某些激素。
血脑屏障的存在对健康至关重要。在阿尔茨海默病等疾病中,覆盖大脑血管的内皮细胞萎缩,有害化学物质得以进入大脑,加重神经损伤。然而,这道屏障也给医学带来了困难——脑癌很难治疗,原因之一就是大多数化疗药物无法穿越血脑屏障抵达肿瘤。
神经元的能量来源
大多数细胞可以利用多种碳水化合物和脂肪作为能量来源,但脊椎动物神经元几乎完全依赖葡萄糖。由于葡萄糖的代谢过程需要氧气,神经元需要稳定的氧气供应,大脑因此消耗了全身约20%的氧气。
神经元之所以对葡萄糖如此依赖,并非因为缺乏代谢其他燃料的能力,而是因为葡萄糖在婴儿期之后实际上是唯一能穿越血脑屏障的主要营养物质——酮体虽然也可以穿越,但正常情况下其数量很少。
大脑只占体重的约2%,却消耗了全身20%的氧气和25%的葡萄糖。这种高度集中的能量消耗反映出神经系统持续运转所需的巨大代谢需求。
葡萄糖的供应通常不会成为问题,因为肝脏可以从多种碳水化合物、氨基酸以及脂肪分解产物中合成葡萄糖。真正可能出现问题的,是葡萄糖无法被正常利用的情况。葡萄糖的代谢需要维生素B1,也就是硫胺素。长期缺乏硫胺素会导致神经元死亡,引发一种称为科萨科夫综合征的疾病,其主要表现是严重的记忆障碍。这种情况在慢性酗酒者中较为常见,因为长期大量饮酒会严重干扰硫胺素的吸收和利用。
理解单个神经元的特性是研究行为的基础,但行为本身并不来自任何一个神经元。就像化学家必须了解原子才能理解化合物一样,神经科学家必须了解细胞才能理解神经系统。然而,神经系统并不是个体细胞的简单叠加,就如同水并不只是氧和氢的简单混合一样。真正产生行为的,是神经元之间的交流与协作。
轴突的信号传递方式
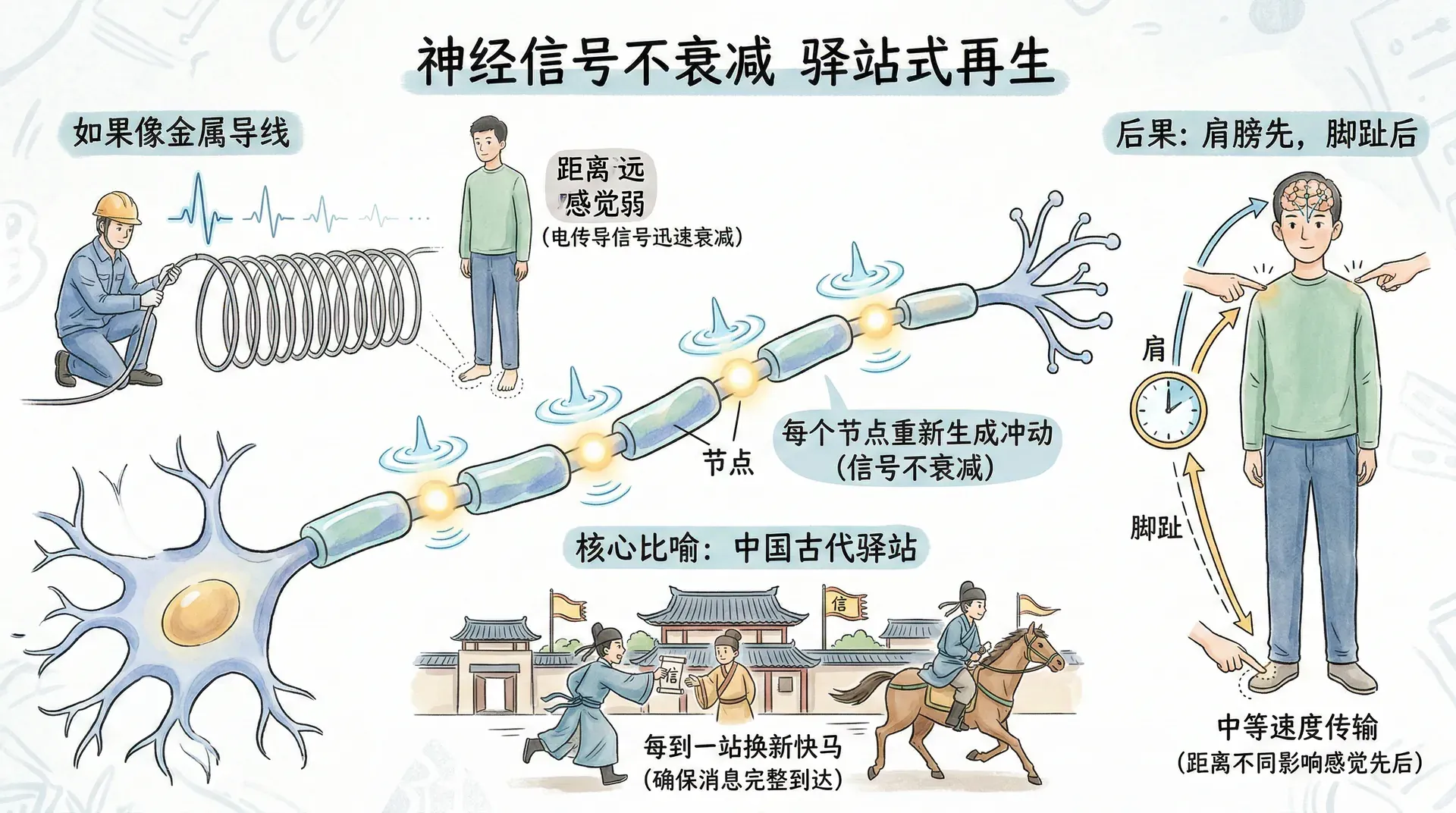
从脚部的触觉感受器到脊髓和大脑,如果轴突像金属导线那样传导电流,冲动强度会在传播过程中迅速衰减。这样一来,身体距大脑越远的部位,感觉就会越弱,矮个子会比高个子更容易感觉到脚趾的存在。这显然与现实不符。
轴突采用了一种避免上述问题的方式:它不是简单地传导电冲动,而是在每个点上重新生成冲动。这就像中国古代的驿站传信,每到一站换新的快马,确保消息能够完整地传到目的地。冲动沿轴突传播而不会衰减,因为每个节点都重新生成了它。
这种方式虽然解决了信号衰减的问题,却带来了另一个细节:由于轴突以中等速度传输信息,肩膀上的触摸会比脚趾上的触摸更早到达大脑。
如果有人同时触摸你的肩膀和脚趾,你并不会注意到两个信号到达大脑的时间差,因为那个差距非常小。实际上,如果有人先后触摸你的两只手,除非间隔超过70毫秒,否则你无法准确判断哪只手先被触摸。
对于触觉而言,大脑并不需要精确记录不同部位信号到达时间的细微差异。但在视觉中情况不同——视网膜上两个相邻点几乎同时发出的冲动,时间上的微小差别就能反映光线的移动方向。为了尽可能准确地检测运动,视觉系统会补偿视网膜不同部位距大脑远近不同的影响:距离更远的部位,对应的轴突传导速度也稍快一些。总体而言,轴突的信号传导特性与神经系统的实际需求高度吻合。
静息电位
神经元中的信息产生于静息电位的变化。静息电位是指神经元在没有受到刺激时,细胞膜内外两侧维持的稳定电压差。
神经元的所有部分都被约8纳米厚的细胞膜覆盖,由两层磷脂分子组成。嵌在磷脂中的是圆柱形蛋白质分子,各种化学物质可以通过这些分子穿过细胞膜。在没有外部干扰的情况下,细胞膜维持着一个稳定的电梯度,即极化状态——细胞内外电荷存在差异。膜内的神经元相对于外部具有略微负的电位,主要原因是细胞内含有大量带负电的蛋白质。这种电压差在典型情况下约为-70毫伏,不同神经元会有所差异。
静息状态下的离子分布
如果带电离子能够自由流过细胞膜,膜就会去极化,静息电位随之消失。然而,细胞膜是选择性透过的,只有特定物质才能通过。氧气、二氧化碳、尿素和水可以自由穿过始终开放的通道,而大多数带电离子则需要通过有时开放有时关闭的膜通道才能穿越。
钠离子和钾离子是调控静息电位的关键离子。当细胞膜处于静息状态时,钠通道关闭,几乎阻止了所有钠离子的流动;钾通道则几乎关闭但不完全关闭,允许少量钾离子缓慢流动。与此同时,钠钾泵作为一种蛋白质复合物,持续将三个钠离子泵出细胞,同时将两个钾离子抽入细胞。这一主动转运过程需要消耗能量,其结果是钠离子在膜外的浓度比膜内高10倍以上,而钾离子在膜内的浓度则比膜外高。
离子所受的两种力
钠离子和钾离子在细胞膜两侧同时受到电梯度和浓度梯度两种力的作用,二者的相对大小决定了离子的净流动方向。
以钠离子为例,在静息状态下,电梯度和浓度梯度都倾向于将钠离子推入细胞:细胞内部带负电,对带正电的钠离子有吸引力;而钠离子在膜外的浓度更高,依据扩散原理,也倾向于向浓度低的细胞内流动。两种力的叠加意味着,一旦钠通道打开,钠离子将迅速涌入。
钾离子的情况则不同。电梯度倾向于将带正电的钾离子拉入细胞,但浓度梯度却倾向于将细胞内浓度更高的钾离子推出细胞。这两种力方向相反,几乎相互抵消,因此静息状态下钾离子的净流动量很小。
静息电位的意义
维持静息电位需要消耗大量能量,身体之所以愿意付出这个代价,是因为静息电位为神经元的快速响应做好了准备。膜通过持续维持钠离子的浓度梯度,相当于提前储存了驱动力——一旦刺激到来,打开钠通道,钠离子便会在两种力的共同作用下快速涌入,产生强烈的电信号。这就像拉满的弓随时准备射出箭矢,等待的过程是能量的积蓄,而不是浪费。
动作电位
轴突发出的信号被称为动作电位。动作电位是一种沿轴突传播的短暂而剧烈的电信号,是神经元在受到足够强烈刺激时,细胞膜电位迅速发生变化的过程。
具体来说,当神经元膜内外的电压差因外部或内部刺激而达到某个临界点(阈值)时,原本相对负电的膜内会在极短时间内变得比膜外更为正电,这一电位的剧烈逆转即为动作电位的产生。
这个过程只持续几毫秒,却足以使信号可靠、高速地从神经元的一端传递到另一端。动作电位的发生标志着神经元“放电”,并将信息以全或无的方式精确传递,是神经系统进行信息交流和调控的基本生物电事件。
阈值与全或无定律
当给神经元施加一个较弱的去极化电流时,膜电位只会略微升高,刺激停止后便恢复原来的静息水平。但当刺激强度超过某个临界点时,情况发生了根本性的变化。这个临界点称为兴奋阈值,一旦膜电位达到阈值,大量钠通道同时打开,钠离子在电梯度和浓度梯度的双重驱动下迅速涌入细胞,膜电位急剧上升,甚至超过零点达到正值,形成一个尖锐的电位峰值,这就是动作电位。
动作电位遵循全或无定律:只要刺激超过阈值,产生的动作电位幅度和速度就基本固定,不会因为刺激更强而变得更大。就像水坝的闸门,达到一定水压才会开闸放水,而水压再大,放出的水量也是由闸门决定的,不会随压力无限增大。
动作电位的分子过程
动作电位的产生和恢复经历了一个有序的分子事件序列。
-
起始状态下,钠离子主要分布在神经元外部,钾离子主要分布在内部,膜处于静息极化状态。
-
当去极化达到阈值时,电压门控钠通道大量打开,钠离子快速涌入,膜电位急剧升高。
-
在电位峰值时,钠通道突然关闭,并在接下来的数毫秒内对重新开放产生抵抗。与此同时,钾通道充分打开,此时细胞内已出现轻微正电荷,浓度梯度和电梯度共同将钾离子推出细胞,带走正电荷,使膜电位迅速回落。
-
由于钾通道在钠通道关闭后仍保持开放,大量钾离子持续外流,使膜电位短暂低于静息水平,形成超极化。随后,钾通道逐渐关闭,膜电位回到正常静息水平。
-
最终,钠钾泵重新建立离子分布,为下一次动作电位做好准备。
局部麻醉药物如普鲁卡因和利多卡因通过与钠通道结合,阻止钠离子进入,从而阻断动作电位的产生。去牙科拔牙时注射的麻醉药正是利用这一原理:感受器感受到疼痛,但轴突无法将信号传送到大脑,疼痛感因此消失。
不应期
动作电位发生后,神经元会进入一段短暂的不应期,在此期间对新的刺激产生抵抗。不应期分为两个阶段:在绝对不应期内,无论施加多强的刺激,膜都不能产生新的动作电位;在随后的相对不应期内,需要比平常更强的刺激才能触发动作电位。这一机制确保了动作电位沿轴突单向传播,同时也限制了神经元发放信号的最高频率。
在大多数神经元中,绝对不应期约为1毫秒,相对不应期约为2到4毫秒。
频率编码
由于全或无定律的限制,轴突无法通过增大单个动作电位的幅度来表示刺激的强弱。它能改变的只是动作电位发放的时间模式。一般规律是,刺激越强,单位时间内产生的动作电位越多,即发放频率越高。除了频率,神经元还可以通过发放节律传达不同类型的信息。味觉轴突对甜味和苦味会显示出不同的发放节律,这是另一种编码信息的方式。
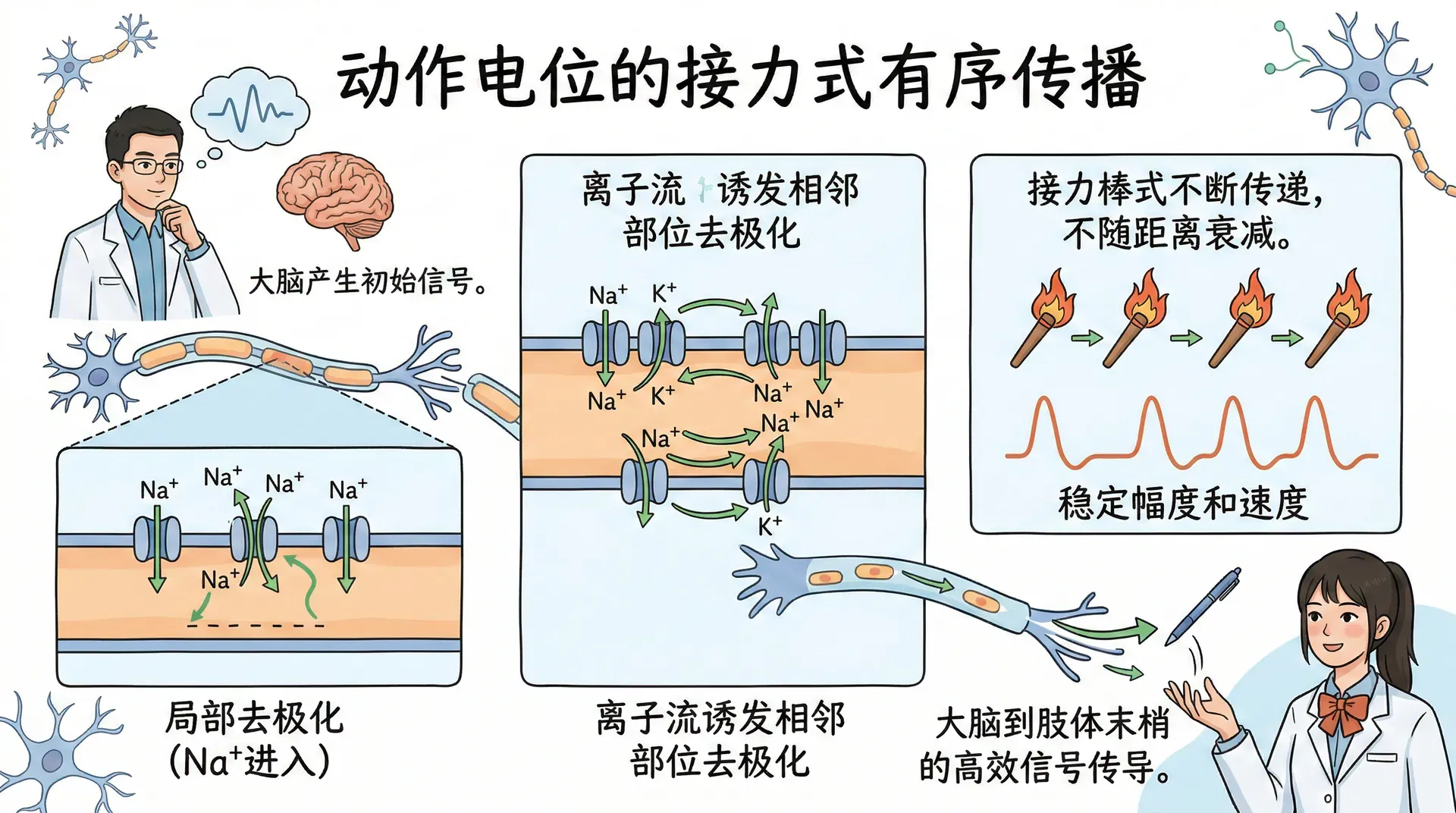
动作电位的传播与髓鞘
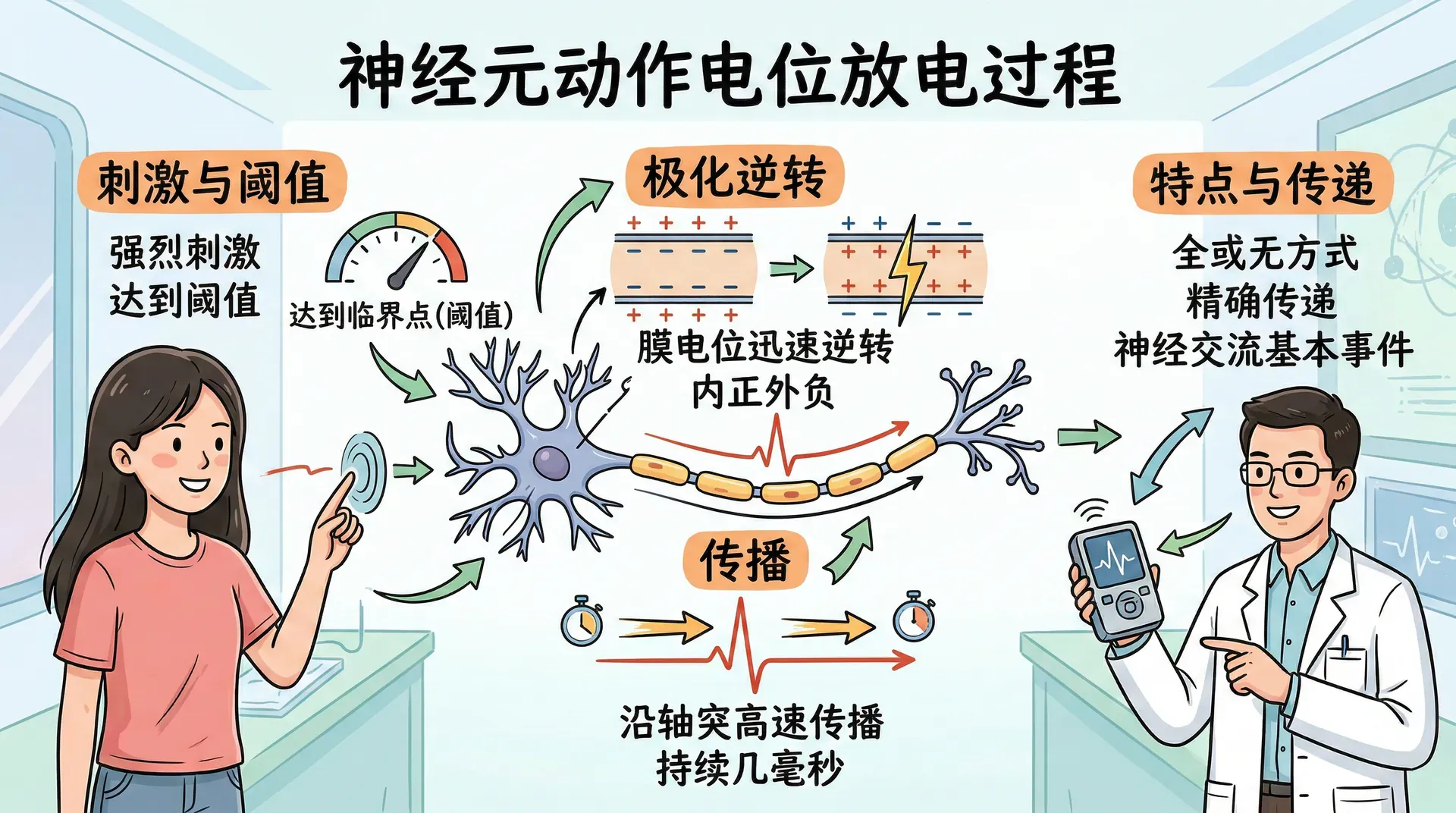
动作电位如何能沿着轴突传播到远离起点的地方,是神经信号能够跨越身体长距离、迅速精确传递的关键。这个机制涉及膜在局部产生的动作电位通过膜内外离子流,诱发相邻部位的去极化,从而使动作电位如接力棒般不断在轴突上传递。无论刺激从哪里开始,动作电位都能以稳定的幅度和速度沿轴突方向有序传播,不会随距离衰减,实现从大脑到肢体末梢的高效信号传导。
传播的基本原理
在运动神经元中,动作电位从轴突小丘——轴突离开胞体的起始部位——开始。当某一点发生动作电位时,大量钠离子涌入,使该点暂时带正电。这些正离子在轴突内部向邻近区域扩散,略微去极化相邻的膜,使其也达到阈值,从而在邻近位置重新生成动作电位。以这种方式,动作电位从一点到下一点逐步传播,信号强度始终不变。
正电荷实际上向两个方向扩散,但动作电位刚刚通过的区域正处于不应期,无法再次被激发,因此信号只能朝前传播,而不会原路返回。
髓鞘与跳跃式传导
在最细的轴突中,动作电位的传导速度不到每秒1米。适当增加轴突直径可以将速度提高到约10米/秒,但这对于需要快速响应的神经系统来说仍然不够。脊椎动物演化出了一种更有效的解决方案:髓鞘。
髓鞘是包裹轴突的绝缘材料,由少突胶质细胞或施万细胞缠绕而成。髓鞘并不连续,每隔一段距离就有一个裸露的小节点,称为郎飞结。由于髓鞘覆盖的区域几乎没有钠通道,动作电位无法在髓鞘下的膜上逐点再生。取而代之的是,离子电流在郎飞结处产生动作电位后,正电荷在轴突内部迅速扩散到下一个郎飞结,在那里重新激发动作电位。这种从结到结的跳跃式传导速度远快于逐点传播,最高可达每秒100米以上。
多发性硬化症是一种免疫系统攻击髓鞘的疾病。髓鞘形成后,被覆盖区域的轴突膜会逐渐失去钠通道。一旦髓鞘被破坏,这些区域因为缺乏钠通道,大多数动作电位都会在两个郎飞结之间消失,无法继续传播。这就是多发性硬化症患者会出现视觉障碍、肌肉协调性差等多种神经症状的原因。
局部神经元
轴突可以产生动作电位并进行远距离传播,但许多小神经元并没有轴突。这类神经元只与距离最近的邻居交换信息,因此被称为局部神经元。由于没有轴突,它们不遵循全或无定律。当局部神经元接收到刺激时,产生的是分级电位——电位变化的大小与刺激强度成正比,并在向周围传播的过程中逐渐衰减。
局部神经元的研究难度较大,因为它们体积极小,插入电极时极易造成损伤。早期神经科学家主要研究大型神经元,对小神经元了解甚少,甚至有人误以为它们是尚未发育成熟的细胞。这种误解很可能是“人类只用了大脑10%”这一说法的来源之一,而这个说法在科学上并不成立。
从神经元到行为
神经元是神经系统功能的基本单元,但行为并不来自任何单一神经元的活动,而是来自数以亿计的神经元之间精密的电化学交流。理解单个神经元的工作原理,是理解整个神经系统乃至人类行为的第一步。
神经系统由神经元和胶质细胞共同构成。神经元负责接收、整合和传递信号;胶质细胞则在维护、保护和支持方面发挥不可替代的作用。血脑屏障保护大脑免受外来有害物质的侵扰,同时通过主动转运机制确保葡萄糖等营养物质的供应。静息电位为神经元储备了随时响应的能量基础,动作电位则以全或无的方式将信号可靠地沿轴突传出,髓鞘的存在使这一过程得以高速且节能地完成。
每一个想法、每一种情绪、每一个动作,背后都是数十亿个神经元之间持续发生的电化学对话。从微小的离子流动到复杂的意识活动,这一跨越尺度的整合过程,正是神经科学最引人入胜的研究领域。
理解这些基本机制不仅具有理论意义,也直接关系到临床医学。局部麻醉药物利用钠通道阻断原理消除疼痛;多发性硬化症的症状根源在于髓鞘损伤;科萨科夫综合征的记忆障碍则源于葡萄糖代谢的受损。对神经细胞运作方式的深入认识,是理解和治疗各类神经系统疾病的重要前提。
“在神经系统这个精密的网络中,每一个细节都蕴含着秘密、神奇和惊喜。”——拉蒙-卡哈尔