神经元
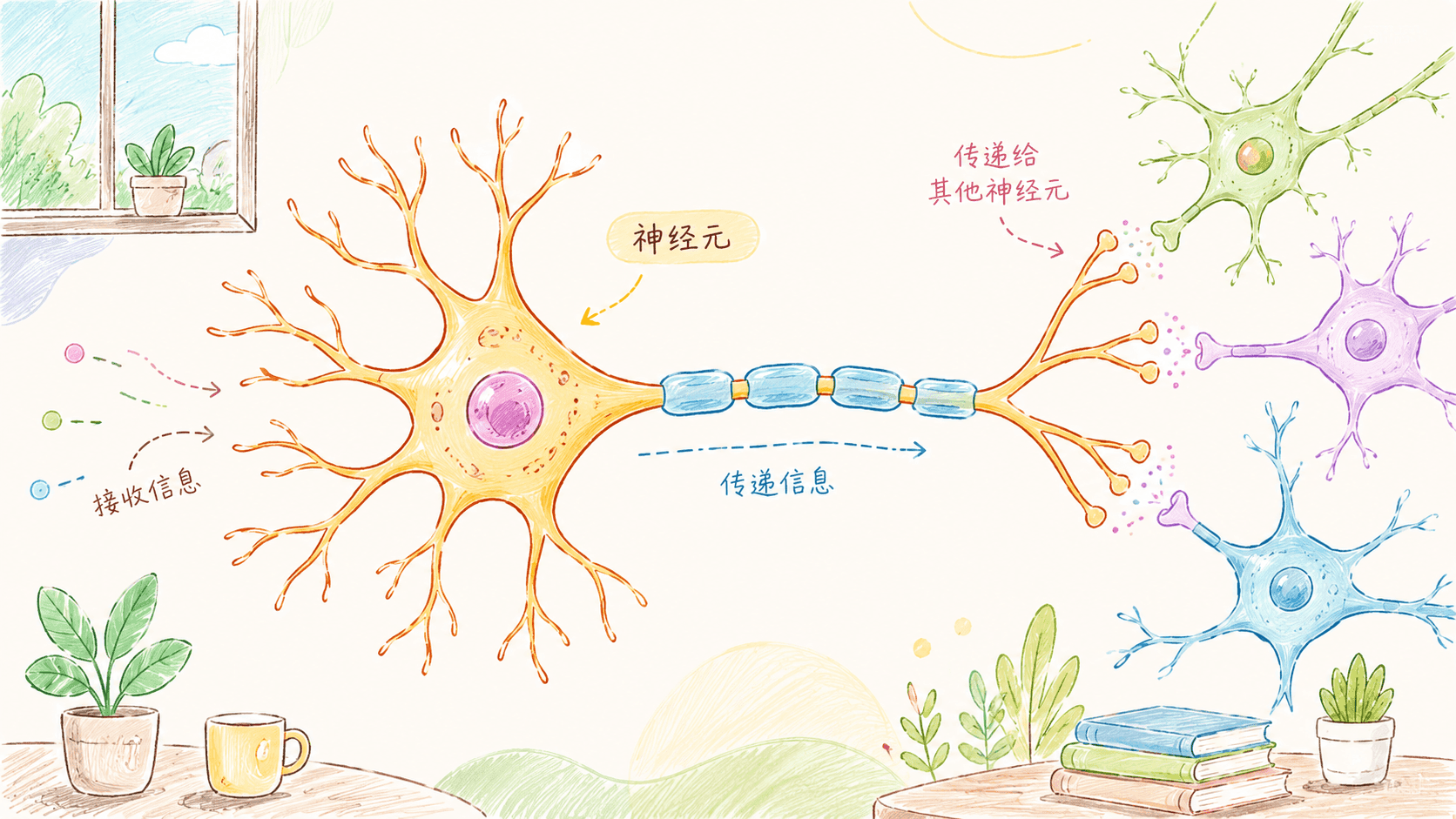
神经元(Neuron)是神经系统不可或缺的基础单元,被誉为“大脑的信息处理器”。每一位人类在出生时,大脑内便拥有数百亿个神经元,这些神经元彼此错综复杂地连接,形成庞大的神经网络。我们的感知、情绪、思考、记忆、决策,乃至每一个身体动作,都离不开神经元之间持续不断的信息传递和协作。
神经元能够感知体内外的各种刺激(例如声音、光线、温度、痛觉),将这些信号转化为快速的电脉冲,并以极高的速度沿着自身的“线路”——轴突传递。同时,神经元之间通过特殊的连接点——突触,将电信号转为化学信号,实现神经元之间的高效“对话”。这种沟通方式保证了我们可以瞬间反应、灵活学习新知识、不断调整行为模式。
神经元的信息传递机制解释了许多我们日常生活中的现象:比如为什么重复练习能让某项技能越来越熟练,是因为相关神经回路的连接被反复强化;又比如药物、激素、甚至情绪变化,都是通过影响神经元之间信号传递或神经递质的分泌,进而影响我们的感觉、心情或行为。深入理解神经元的结构与运作,是认识人类大脑、心理及行为机制的基础,也是现代神经科学研究的核心起点。
神经元的基本结构
人脑中约含有860亿个神经元,但这只占整个大脑细胞总数的一小部分。实际上,大脑内存在数量与神经元大致相当的“胶质细胞”——这些细胞在过去曾被误认为只是“神经系统的胶水”,但现代神经科学发现,它们不仅为神经元提供结构性支持、营养、绝缘与清除代谢废物,还主动参与调节神经元间的信息传递。没有胶质细胞的协作,神经元无法长期独立生存和正常发挥功能,二者共同维系着神经系统的高效运行。
神经元和一般身体细胞最大的不同,在于其复杂而高度专化的结构,使其能够敏锐地感知环境、快速传递和处理大量信息。一个典型的神经元主要由以下三大部分组成:细胞体(soma)、树突(dendrites)、轴突(axon),每一部分各司其职、紧密协作,支撑着神经系统的整体功能。
细胞体(Soma)是神经元的“司令部”,也是代谢和生命活动的主控中心。细胞体内包含细胞核,存储着完整的DNA用于遗传信息的表达与蛋白质合成;丰富的线粒体为能量消耗极高的神经元不断供能。此外还有高尔基体、内质网等各种细胞器,共同维系细胞的基本运作。细胞体的另一大任务,是对树突接收到的所有电化学信号进行整合和处理——只有当这些信号的总和达到一定门槛时,才会触发新的动作电位沿轴突传递下去。
树突(Dendrites)则像树木繁复的枝丫,从细胞体四周向外扩展,面积广阔,是神经元最主要的“接收天线”。每条树突表面密布着上千个受体蛋白,能够响应来自其他神经元的神经递质信号。通过这些“接收器”,一个神经元可以与成千上万个其他神经元建立突触连接,实时获取来自体内外环境的多样化信息。树突的分布和结构形态极为多样,正是这种复杂的分支结构,使神经元具备了高维度的信息集成与调控能力。
轴突(Axon)是神经元的“信息高速公路”,呈细长的纤维状,从细胞体只延伸出一根主干,可以短至几毫米,也可长达一米以上(如控制下肢运动的运动神经元),其长度和直径与功能密切相关。轴突最重要的作用,是将细胞体整合后的“决策信号”快速、远距离地传递给下一个神经元或效应器(如肌肉、腺体)。很多轴突外被一层或多层髓鞘包裹,大大提升信号传导速度与可靠性。轴突末端常分支扩展,形成“轴突终末”,在此通过突触结构与下游神经元发生连接,实现进一步的信息传递。
此外,神经元间的连接千变万化:有的神经元树突密集、能够处理复杂信息,有的轴突极长、负责远距离传递指令,有的则参与形成局部微型神经回路。这种结构多样性和高度分化,是大脑具备惊人计算能力和灵活性的根本保障。
神经元的具体形态千差万别,例如锥体细胞、星形细胞、篮细胞等均有自己独特的结构特征和功能分工,这种结构与功能的精细对应,使大脑的不同区域能够处理各类复杂任务。
电信号是如何产生的
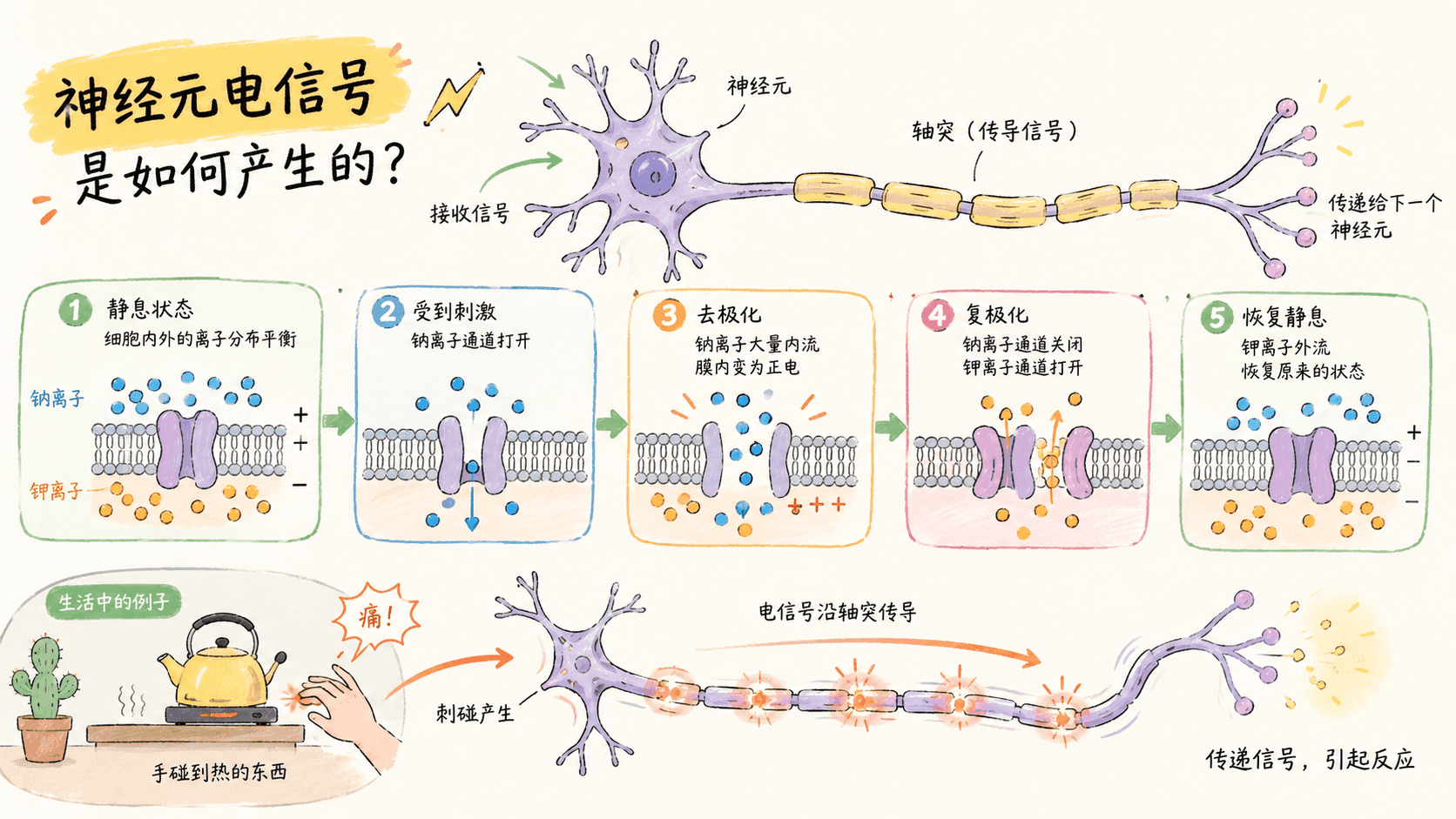
神经元之所以能够进行信息传递,核心就在于一种被称为“动作电位”(Action Potential)的电信号。那么,这个电信号是如何产生和传播的呢?首先,我们需要理解神经元细胞膜内外的“离子浓度差”——这是一切电信号的源头。
在静息状态下,神经元细胞膜内侧含有较高浓度的钾离子(K⁺),而膜外则富含钠离子(Na⁺)。这种离子分布并非偶然,而是依赖于膜上的钠钾泵等特殊蛋白主动运输维持的。正因如此,神经元膜两侧形成了一个稳定的电位差,静息状态下一般为-70毫伏(mV),即细胞内电位比外侧负得多,这被称为“静息电位”。
当神经元的树突或细胞体受到刺激时,膜电位会发生一系列变化,这一过程可分为以下几个关键步骤:
受刺激:树突或细胞体受到外界或其他神经元信号刺激,膜电位开始改变。
达到阈值:如果刺激足够强,使膜电位提升到大约-55毫伏(称为“阈值”),则触发下一步反应。
钠离子通道开放:膜上的钠离子(Na⁺)通道在瞬间全部打开,大量Na⁺迅速进入细胞内,使膜电位由负转正,通常会上升到约+30毫伏。
钾离子通道开放:随后,钾离子(K⁺)通道也随之开放,K⁺流出细胞,膜电位迅速恢复到静息水平。
形成动作电位:上述电位的剧烈变化就是“动作电位”(神经冲动)。整个过程仅需1-2毫秒,非常迅速。
动作电位的产生拥有“全或无”的特性——要么不产生,要么就产生一次标准幅度、不可“打折扣”的动作电位。因此,神经元不会因为刺激轻微就产生微弱的动作电位。不同强度的信号是通过单位时间内动作电位发放的“频率”编码的:微弱触觉只会让神经元慢慢发放,而强烈刺激则会大大提高动作电位产生的频率,最多可达每秒几百次。这种“频率编码”使大脑得以区分不同强度的感受。
值得一提的是,动作电位沿着轴突的传导速度并不是固定不变的,而是与轴突的结构密切相关。没有髓鞘包裹的裸露轴突,动作电位的传播速度只有0.5 ~ 2米/秒;而有髓鞘的轴突,通过“跳跃式传导”(动作电位在各朗飞结之间跳跃),传导速度可以高达70 ~ 120米/秒,足足快了60倍以上。这就是为什么有些反应可以极其迅速,而某些慢痛传导却较为迟缓。
髓鞘
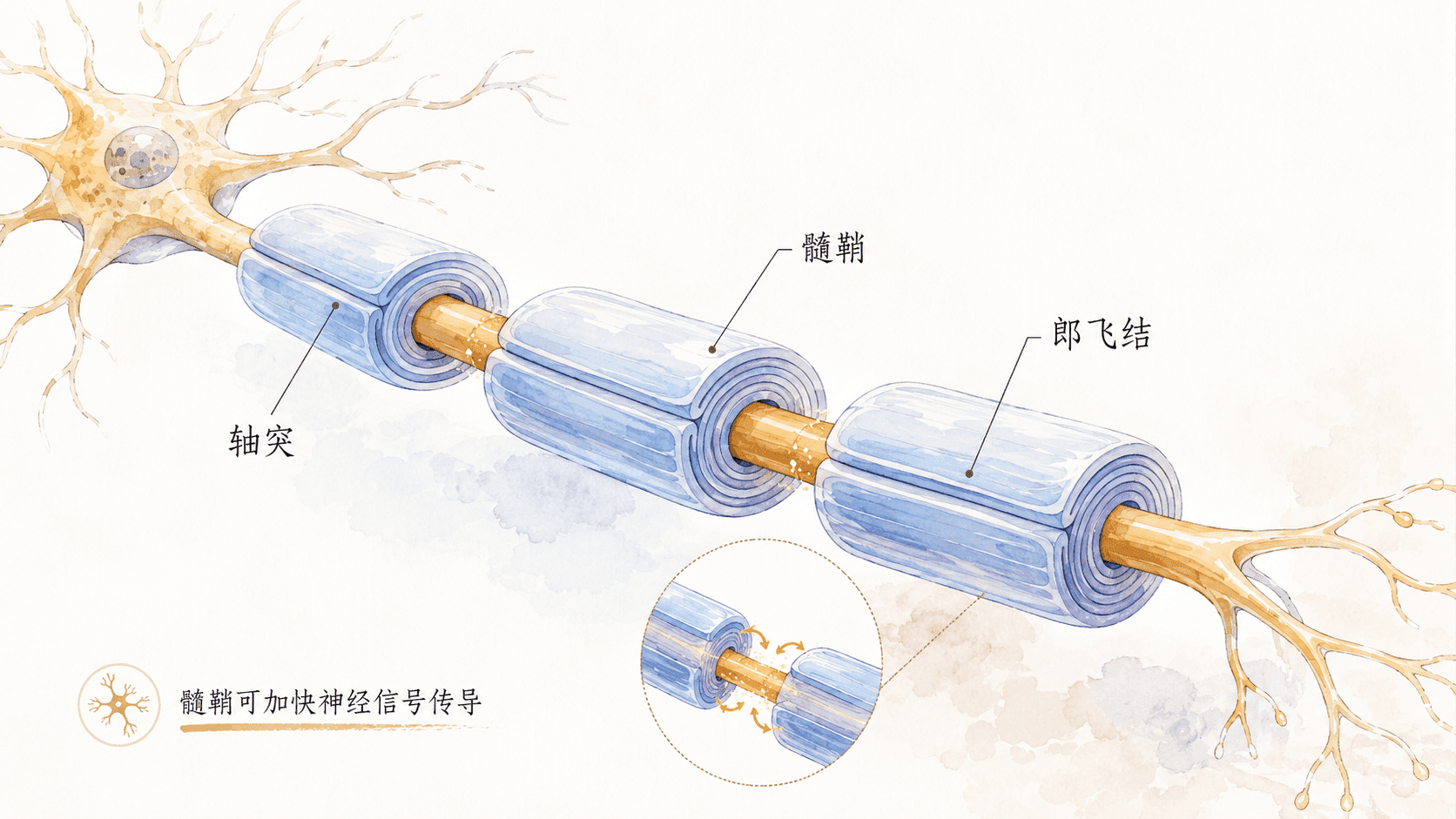
在许多神经元的轴突外部,都会包裹着一层由脂质和蛋白质构成的绝缘鞘,称为“髓鞘”(Myelin Sheath)。髓鞘好比是电线上的绝缘皮层,能有效防止电信号泄露,并提高神经冲动传递的速度。值得注意的是,髓鞘并不是沿轴突连续包裹,而是呈分段结构,每两段髓鞘之间留有一个暴露的空隙,被称为“朗飞结”(Node of Ranvier)。
当动作电位顺着有髓鞘的轴突传导时,并不需要沿着轴突缓慢地逐点扩散,而是以“跳跃式”的方式,从一个朗飞结快速跃迁至下一个,这种机制大大地提升了信号传输效率,同时也能显著降低能量的消耗,对机体节约神经能量有重要价值。
髓鞘的生理意义极为重要,主要包括:
- 在脊髓和大脑等中枢神经系统中,髓鞘由“少突胶质细胞”(Oligodendrocyte)产生;
- 在周围神经系统中,髓鞘主要由“施万细胞”(Schwann cell)构成;
- 髓鞘的生成过程漫长复杂,从胎儿期就已开始,在出生后快速推进,直到青少年晚期(约25岁)才完全完成;
- 大脑额叶的髓鞘化发育最晚,这解释了青春期及青年早期冲动控制能力仍不成熟的神经生物学基础;
- 髓鞘不仅有助于实现高速神经传导,还对神经元的生存与健康具有养护作用。
临床上,与髓鞘相关的疾病极为典型。以多发性硬化症(MS, Multiple Sclerosis)为例,这是一类自身免疫疾病——患者的免疫细胞错误地攻击并损伤中枢神经系统的髓鞘,导致神经信号的传导变慢、失真甚至完全中断。这会引起一系列多样化的神经症状,如肌肉无力、感觉障碍、行动协调失常、视力下降等。不同病人的症状表现会因髓鞘受损的区域和严重程度各不相同。
下方总结了不同神经纤维髓鞘化状况与传导速度、功能的对应关系:
突触
神经元之间的联系并非像电线一样直接物理连接,而是在相邻两个神经元之间存在一个极其微小的空隙,称为“突触间隙”(synaptic cleft),其宽度一般仅约为20纳米,相当于头发丝直径的五千分之一。突触是神经信号交流的枢纽,也是神经信号由电信号与化学信号之间双向转换的关键场所。
神经信号跨越突触的主要过程如下:当动作电位沿轴突传递到达轴突末端时,会促使“突触囊泡”中的神经递质释放到突触间隙。随后,神经递质分子迅速扩散并与下一个神经元(突触后神经元)树突或细胞体膜上的特定受体结合。这一结合会导致突触后膜发生电位变化,表现为兴奋性或抑制性的反应,从而影响该神经元是否产生新的动作电位。
更精细地说,不同的神经递质与不同的受体结合,会激发截然不同的生理效应,主要包括:
- 谷氨酸(Glutamate):最主要的兴奋性神经递质。能够使突触后膜电位向上变化,使神经元更容易产生新的神经冲动。
- GABA(γ-氨基丁酸):主要的抑制性神经递质。能够使突触后膜电位下降,从而抑制动作电位的发放。
每个神经元在某一时刻是否被“点燃”,取决于它同时接收到的所有突触兴奋性输入与抑制性输入的总和。这种复杂的同步整合能力赋予神经网络高度可塑性。
在神经递质完成其信号传递使命后,必须迅速被清除以保证信号的分辨率和神经活动的准确调控。清除机制主要包括:递质分子被突触前神经元重新摄取(再摄取机制,是许多抗抑郁药重要的药理靶点);被特定酶降解分解;或自然扩散离开突触间隙。这些机制确保了神经系统的信号传递既迅速又精确、适时终止,为大脑的复杂活动提供了基础保障。
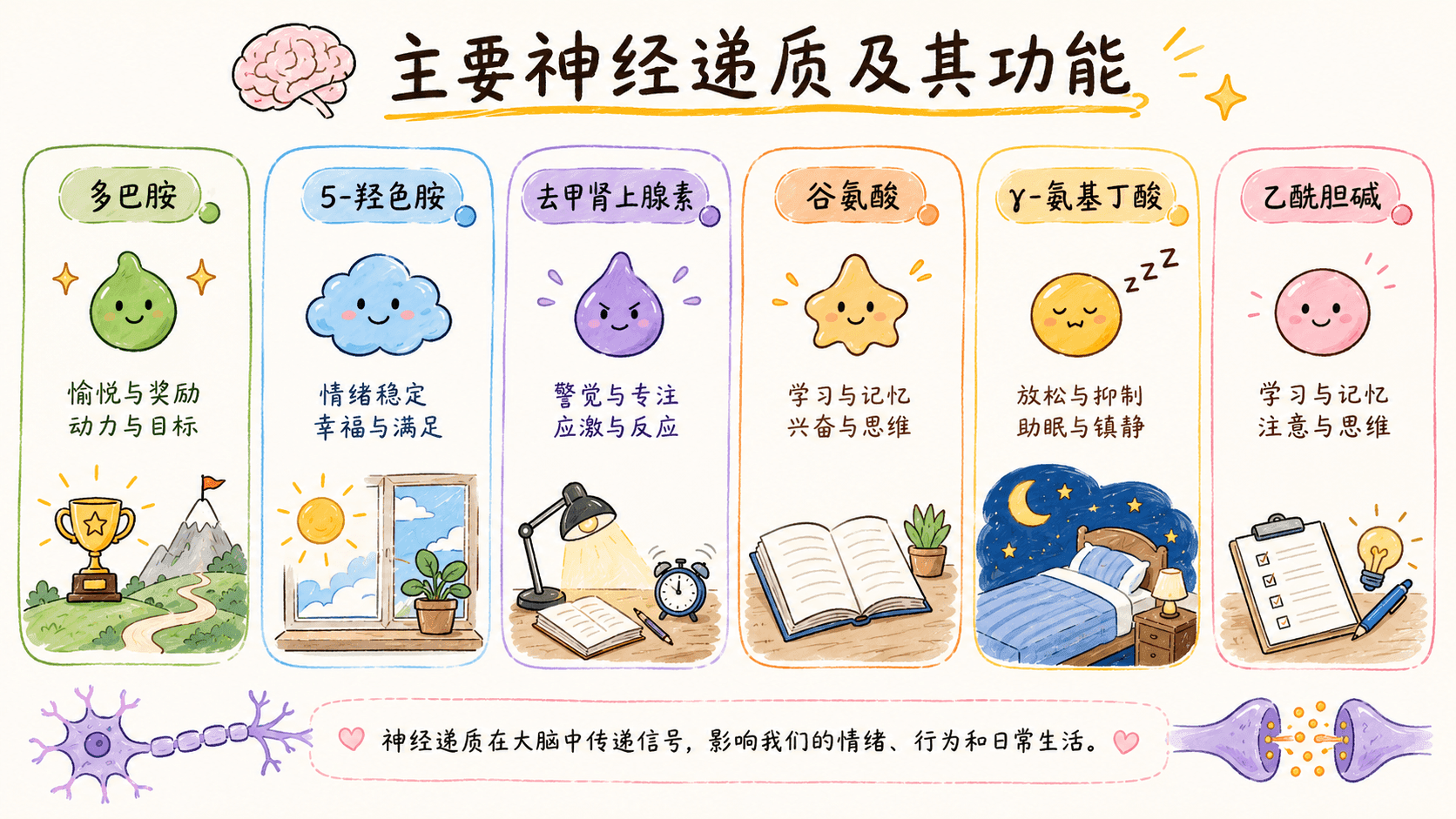
主要神经递质及其功能
目前科学界已知的人体大脑神经递质数量超过100种,这些化学信使物质使神经元之间可以高效地交流信息,参与调控我们的情绪、认知、行为和身体功能。但在如此庞大的神经递质家族中,真正决定日常心理健康、学习记忆和情绪调节的,却集中在少数几种“主角”身上。掌握这些主要神经递质的作用机制,不仅有助于理解药物和疾病如何影响大脑,也能帮助我们更好地认识自己的情绪和大脑运作原理。
谷氨酸(Glutamate)是大脑中含量最高、作用最为广泛的兴奋性神经递质。绝大多数的神经元之间的信息快速传递都依赖谷氨酸完成。特别是在学习和记忆过程中,谷氨酸受体(如NMDA受体、AMPA受体)的激活对于“长时程增强”(LTP,一种突触传递效率提升的机制)的形成至关重要。如果谷氨酸释放失去控制,过度兴奋还可能造成神经元损伤,即所谓的“兴奋性毒性”。
GABA(γ-氨基丁酸,Gamma-aminobutyric acid)则作为主要的抑制性神经递质,扮演着“神经系统刹车手”的角色。它通过降低神经元的兴奋性,防止神经活动失控,有助于大脑整体活动平衡。如果GABA功能减弱,会引发焦虑、易激惹,甚至癫痫。临床上很多抗焦虑、镇静和抗癫痫药物(如苯二氮䓬类安定、咪达唑仑等),正是通过增强GABA的作用来达到疗效的。
多巴胺(Dopamine)是一种调节性神经递质,与奖励机制、动机驱动力、愉悦感、专注力以及运动协调密切相关。比如,当我们完成目标或获得期待的奖励时,多巴胺的分泌量会增加。帕金森病就是由于多巴胺能神经元退化引起的运动障碍;而多巴胺的异常升高则可能导致精神分裂症等精神症状。多巴胺系统在上瘾、情感障碍以及注意力障碍等疾病机制中也占据重要地位。
血清素(Serotonin,5-羟色胺,5-HT)主要负责调节情绪稳定性、睡眠节律、食欲和社交行为。血清素不足和抑郁、焦虑、睡眠障碍密切相关。常见的抗抑郁药(如SSRIs选择性血清素再摄取抑制剂)就是通过提高突触间隙血清素浓度,让情绪趋于稳定和积极。
乙酰胆碱(Acetylcholine)主要涉及学习和记忆能力,参与注意力保持、认知处理和肌肉收缩。阿尔茨海默症患者体内相关胆碱能神经元大幅减少,是其记忆衰退的神经基础之一。部分治疗阿尔茨海默症的药物(胆碱酯酶抑制剂)正是通过延缓乙酰胆碱分解,暂时改善记忆力和认知功能。
去甲肾上腺素(Norepinephrine,又称正肾上腺素)则主管注意力、警觉性、应激和唤醒反应。身体面对压力、危险或刺激时,去甲肾上腺素会让心跳加快、瞳孔放大、注意力高度集中。不足可能导致情绪低落、注意力障碍,而过多则与焦虑、惊恐有关。
这些主要神经递质的类型、功能及其失衡状态与多种常见心理和神经疾病密切相关,可通过下方交互面板逐一深入了解:
除上述外,还有内啡肽(调节疼痛与快感)、组胺(参与觉醒和免疫)等活跃在特定脑区。新型神经递质的发现不断丰富我们对大脑复杂性的理解。
神经可塑性的分子基础
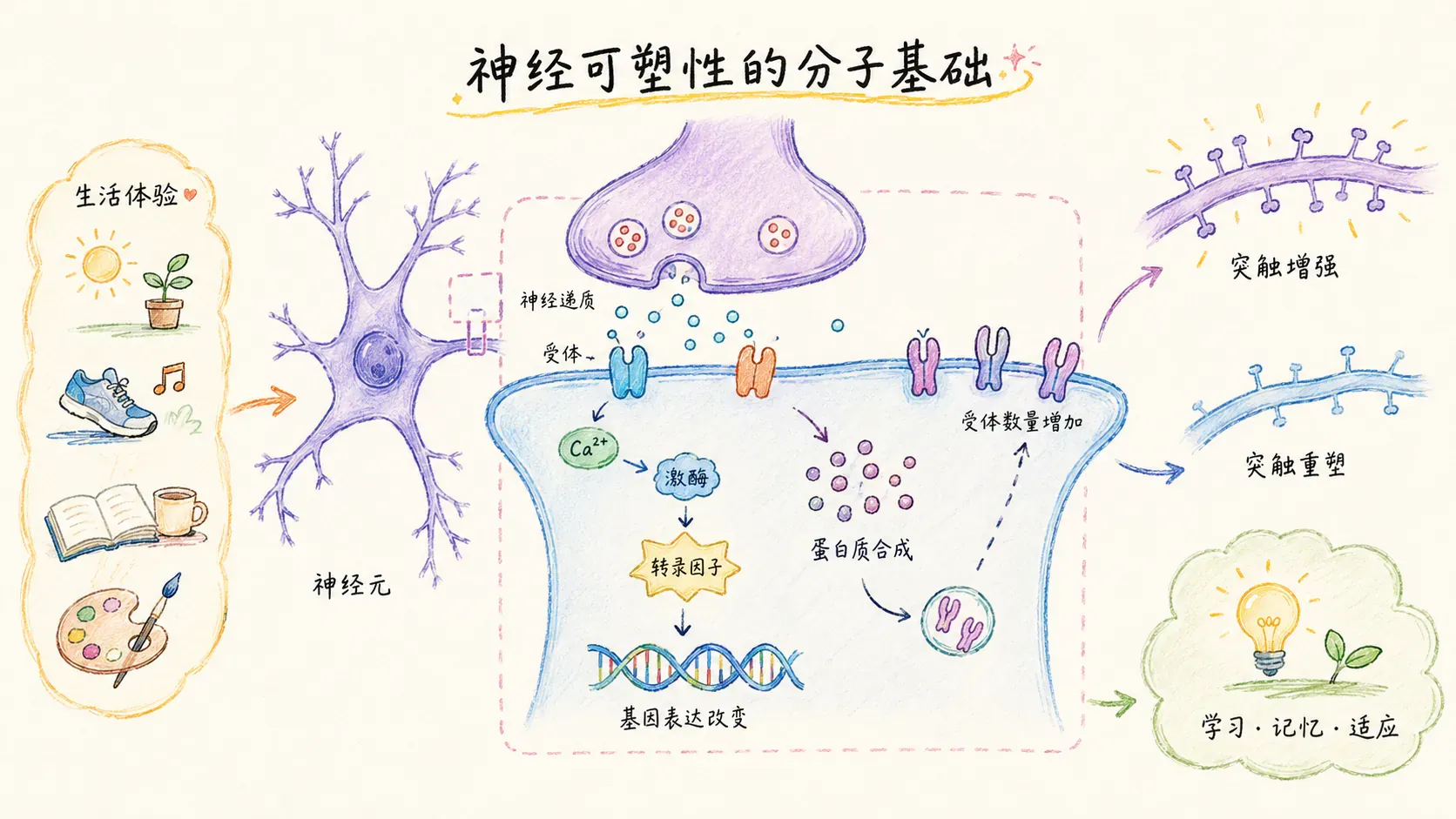
我们所说的“大脑能够改变自己”,其实现机制的核心在于神经可塑性(Neural Plasticity)这一概念。
大脑有一条极为重要的原则,最早由著名神经科学家唐纳德·赫布(Donald Hebb)在1949年提出:“一起激活的神经元,连接在一起”(Neurons that fire together, wire together)。
这句话揭示了一种基本的神经生物学规律:当两个神经元反复同时被激活,它们之间的突触连接就会逐渐加强,信号的传递效率也会提升。这一现象被称作“长时程增强”(Long-Term Potentiation,缩写为LTP)。
LTP(长时程增强)是现代神经科学用来解释学习和记忆为何可以在大脑中持久存储的分子基础。它的关键机制可以归纳如下:
- 反复练习某项技能、反复记忆知识点或多次经历某种体验时,相关神经元之间的突触连接会不断加强,就像把大脑中的“高速公路”加粗加固。
- 具体表现为:突触结构变得更粗壮,神经信号传递更高效。
- 例如,熟练钢琴家的大脑中,负责指法灵活的神经回路,由于高频率练习,突触连接更加稳固、更易激活。
- 这些神经回路结构上的不同,在大脑成像和电生理实验中都能被实际观测和验证。
相反,长期不用或极少激活的突触连接会逐渐削弱,甚至彻底消失,这一过程被称为“突触修剪”。青少年时期是突触修剪高度活跃的阶段——不常用的连接会被大量删除,而经常被激活的线路则不断巩固。这就是“用进废退”原则在大脑结构层面的直接体现:努力和习惯的积累,真的能够改变你的大脑结构。
实际上,神经可塑性的分子机制非常复杂,涉及多种受体(诸如NMDA受体、AMPA受体),信号通路,以及突触结构蛋白的变化等内容。不仅仅是突触强度的增强,突触的数量、形态和分布也会因经验和环境刺激而发生可逆性变化。这种高度动态的可塑性,为我们终身学习和适应新环境提供了可能。
深入理解神经可塑性,对于科学安排学习策略有直接且实用的指导意义:比如“间隔重复”(隔天、隔周反复练习)远比短时间刷题过量(集中突击)更容易让LTP牢固建立;主动回忆(不看笔记试着复述重点、讲给别人听)比单纯重复阅读,更能激活目标神经回路,有效强化突触连接。合理利用可塑性原理,每个人都可以让大脑变得越来越“聪明”。
练习题
第一题
知识点:神经元的基本结构
以下关于神经元结构的描述,哪一项是正确的?
A. 树突负责向其他神经元传出信号,轴突负责接收信号
B. 细胞体是信号传出的出口,轴突是信号整合的中心
C. 树突负责接收来自其他神经元的信号,轴突负责将信号向远端传递
D. 轴突终末直接与下一个神经元的细胞核相连,形成直接电气连接
答案:C
神经元的信息流向是单向的:树突(接收信号)→ 细胞体(整合信号)→ 轴突(传递信号)→ 轴突终末(释放神经递质)。选项A把树突和轴突的功能颠倒了;选项B把细胞体和轴突的角色混淆了;选项D错误,轴突终末与下一个神经元之间存在突触间隙,通过化学信号(神经递质)而非直接电气连接来传递信号。
第二题
知识点:动作电位与“全或无定律”
以下关于动作电位的描述,哪一项是正确的?
A. 动作电位的幅度随刺激强度的增加而增大,强刺激产生更大的电位变化
B. 神经元通过调整单次动作电位的幅度来区分信号强弱
C. 动作电位遵循“全或无定律”,信号强度的差异通过发放频率而非幅度来体现
D. 动作电位只能沿轴突单向传导,如果没有髓鞘则无法传导
答案:C
动作电位遵循“全或无定律”:膜电位达到阈值(约-55毫伏)时,触发标准幅度(约100毫伏)的完整动作电位;低于阈值则不触发。信号的强弱不是通过改变单次电位的幅度,而是通过改变单位时间内发放动作电位的频率(发放率)来编码。选项A、B描述的是错误的幅度调制机制。选项D错误,无髓鞘轴突同样可以传导动作电位,只是速度较慢(0.5-2 m/s),有髓鞘轴突通过跳跃式传导大幅提速。
第三题
知识点:髓鞘的功能与多发性硬化症
多发性硬化症患者出现肌肉无力、感觉异常、协调障碍等多样化症状,根本原因是?
A. 大脑神经元细胞体大量死亡,导致信号无法产生
B. 免疫系统攻击并破坏了神经元的髓鞘,导致神经信号传导减慢或中断
C. 突触间隙中的神经递质被完全耗竭,信号无法跨越突触传递
D. 脑脊液循环障碍,导致大脑营养供应不足
答案:B
多发性硬化症是一种自身免疫性疾病,特征是免疫系统错误攻击中枢神经系统的髓鞘(由少突胶质细胞形成)。髓鞘受损后,轴突的跳跃式传导被破坏,信号传导速度大幅下降或完全中断,导致各种神经系统功能障碍。症状的多样性取决于髓鞘损伤发生在哪些部位。选项A描述的是更严重的神经退行性损伤;选项C、D均不是MS的发病机制。
第四题
知识点:突触传递与神经递质
以下关于突触信号传递过程的描述,顺序正确的是?
A. 神经递质释放 → 动作电位到达轴突终末 → 突触囊泡与膜融合 → 受体结合
B. 动作电位到达轴突终末 → 突触囊泡与膜融合 → 神经递质释放到突触间隙 → 与受体结合触发新信号
C. 树突受体结合神经递质 → 突触囊泡释放 → 动作电位产生 → 信号传向下一神经元
D. 动作电位到达树突 → 直接跨越突触间隙 → 在下一神经元产生新动作电位
答案:B
突触信号传递的标准流程是:动作电位沿轴突到达轴突终末 → Ca²⁺内流触发突触囊泡与突触前膜融合 → 神经递质被释放到突触间隙 → 神经递质扩散并与突触后膜的特异性受体结合 → 引起离子通道开放,改变突触后神经元的膜电位 → 若整合后的电位变化达到阈值,产生新的动作电位。选项A顺序颠倒;选项C逻辑混乱;选项D错误,动作电位到达树突而非轴突,且突触间隙不允许电信号直接穿越,需要化学信使。
第五题
知识点:赫布定律与神经可塑性
根据赫布定律(Neurons that fire together, wire together),以下哪种学习行为最能有效强化神经元之间的突触连接?
A. 把同一段内容连续阅读20遍,每次间隔不超过5分钟
B. 阅读学习内容后立即合上书本,用自己的话尝试复述,并在1天后、1周后各复习一次
C. 将所有学习任务集中在考试前一晚完成,保证单次学习时间足够长
D. 只在感兴趣时学习,不感兴趣时不强迫,以免产生负面联想
答案:B
赫布定律强调的是“反复同时激活”——间隔重复学习能让参与记忆的神经回路在不同时间点多次被激活,每次激活都进一步强化突触连接(LTP)。主动回忆(合上书复述)比被动重读更能有力地激活目标回路。选项A的集中重复缺乏间隔,LTP稳固效果差;选项C的“突击复习”虽然能形成短期记忆,但缺乏间隔激活,长期保留率低;选项D忽视了重复激活对突触强化的必要性。