生物变异
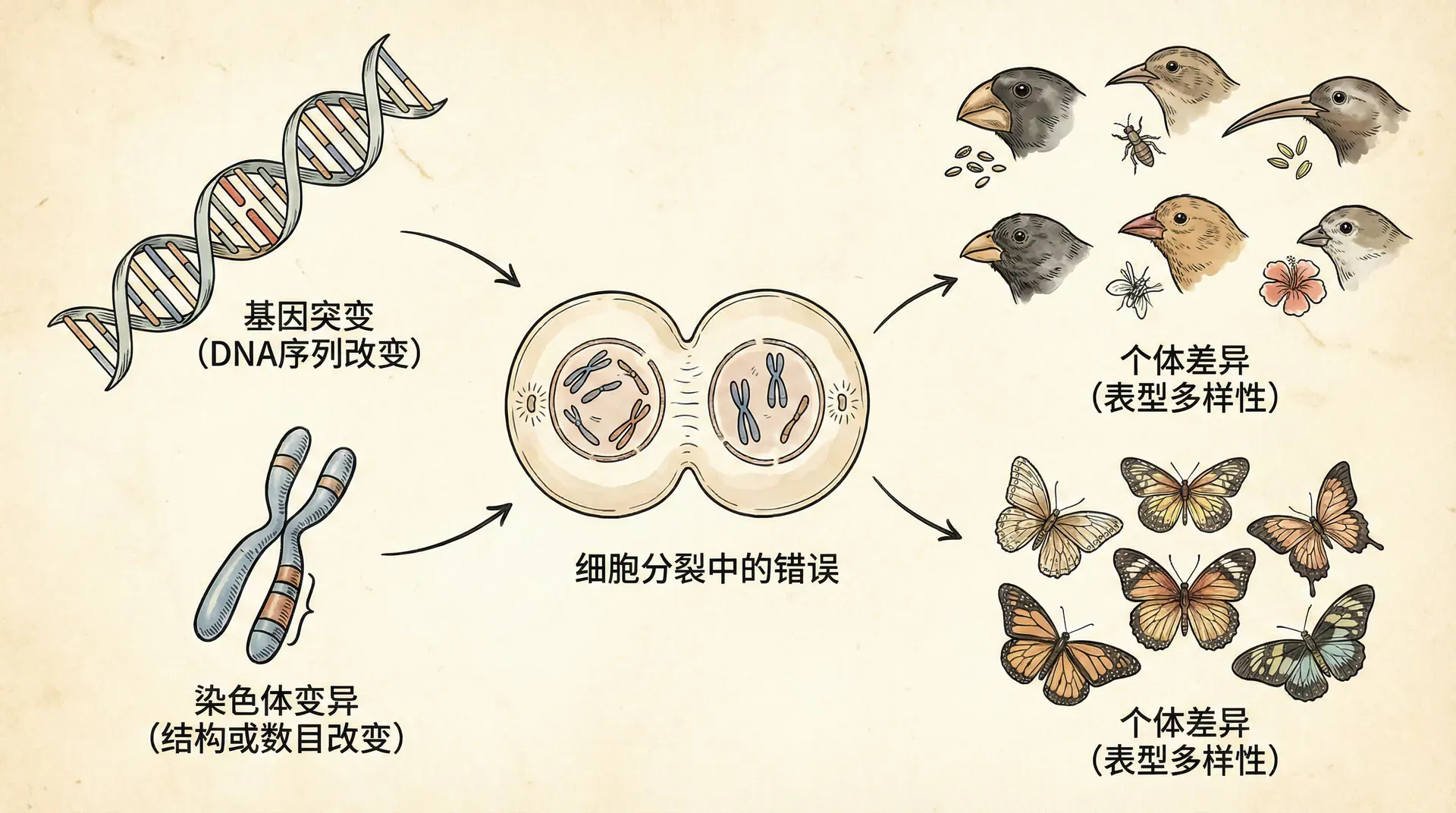
在生命科学的探索过程中,最令科学家们着迷的一件事,是生命体在代际传递中的既有规律性又有变异性。每一代生物在外观、行为乃至细胞层面上,大体都复刻了父辈的特征,但细看之下,却总会发现无数细微乃至偶尔明显的不同点。这些差异——有的微小至难以察觉,有的却足以引发巨大变化——正是驱动物种多样化、推动自然选择和进化过程的主要动力。
如果遗传信息始终无误地复刻,每一代生物都是上一次的完美“复制品”,那整个生物界的适应能力、进化潜力与生态系统的丰富性都将陷入停滞。实际上,正是因为基因复制过程中偶发的变异与环境因素的共同作用,使得每个个体无论在形态、生理还是行为上都可能与祖先略有差异。这些“看似偶然”的不同,不仅为物种应对环境变化提供了原材料,也为科学家们研究遗传机制和进化规律带来了无穷的线索与启发。
变异的“跳跃”现象
传统认知的瓶颈
过去,科学家常常假定所有个体之间的变异都是缓慢、微小且连续地累积,认为这些渐变正是自然选择可用的“原材料”。这种观点在达尔文时代尤为盛行,人们相信无论在多么纯粹的群体内部,总是会自发地产生些许细小而随机的偏差。然而,随着遗传学的发展,这种想法逐渐暴露出局限性。现代研究显示,微小的、分布于某一性状上的连续变化,很多时候都不会被稳定地传递下去,也就是说,这些变化不具备遗传性。
我们用一个实验更直观地感受这一原理:比如选种纯种的东北粳稻,对每穗稻长短作统计,结果多半呈现一个钟型的分布图。不同稻穗围绕着某个平均长度上下浮动,这种浮动看似有规律。也许你会想:如果只留下最长、最理想的稻穗,用它们的种子培育下一代,那么“好”的性状就会被积累,平均稻穗也会变长。
但事实是,如果所用的是遗传上高度纯合的品系,经过这样人为“优选”后的后代表现却几乎不会变,分布图曲线还是原样。即便你只播种最短稻穗的种子,结局一样。“挑选”在这里几乎不产生影响。
这背后真正的原因在于:这些性状的微小波动,大多是环境、营养等偶然因素作用的结果,而不是由可遗传的基因变异造成的。因此,这类连续微小的差异即便再明显,也不会代代相传。
所以说,遗传并不是把所有细节一丝不苟地传递,而是有其内在规律的。只有真正的遗传变异,才可能影响群体后代结构。
从突变到新理论
那么,遗传物质真正的变异会以什么方式呈现?到了20世纪初,德弗里斯通过大量实验发现了令人惊讶的现象:即使在保持种群高度纯合的条件下,偶尔也会冒出极个别的“异类”——这些个体与同伴存在明确的差别,而这种差别不是逐渐积累的,而是突然出现的。德弗里斯将之命名为“突变”。这里的“跳跃”,其实就是指变化的不可连续性——它不经由缓慢的中间态。
突变最大特征是其断层式的不连续,像是某种“质变”的发生。这种概念实际上与当代物理学中的量子跃迁有相似之处。就像电子只能跳到下一个能级、永远不会“半途而废”,遗传的突变也是如此,表现为从一个状态“跳”到另一个全新而确定的状态。
事实上,今天我们已知,分子层面的突变常常涉及到DNA的单点改变、片段置换等机制,这些变化很多都符合“量子跳跃”的离散特征。当然,在德弗里斯提出这些设想时,量子物理还只是刚刚出现的理论前沿,这种跨学科的联想也说明科学的发展总在不断打通新视角。
突变的遗传特性
完全遗传性
突变具有很高的遗传稳定性,这一点与原始、未发生变异的性状完全一致。以之前提到的水稻作物为例,在高度纯合的水稻田中,仍有可能偶尔见到某些稻穗,其长度或者外观特征(如完全没有芒)大幅度超出了原有群体的范围。如果这些表现是由德弗里斯意义上的突变导致的,那么这些变异类型将会以极高的忠实度完全传递给下一代——也就是说,这些无芒的稻穗,其所有后代也都将保持无芒的性状。这一现象在植物、动物和微生物中普遍成立,如我们在园艺作物、实验小鼠和细菌的单克隆培养中都可以直接观察到类似例子。
突变是遗传库中产生创新性状和新变异的根本动力。每一次突变,实际上就是“遗传财富”的一次真正改变,而这种改变只能通过遗传物质本身的原子级结构变化来解释。大多数现代育种实验、遗传学分析,以及植物和动物新品种的选育,基本思路都是通过有计划地杂交,把突变个体(或携带多重突变的个体)与不同类型的个体交配,从而观察、追踪突变性状的遗传分布和表达规律。有时还利用分子标记或基因组测序,精确锁定突变发生的基因位点。
遗传模式的理解
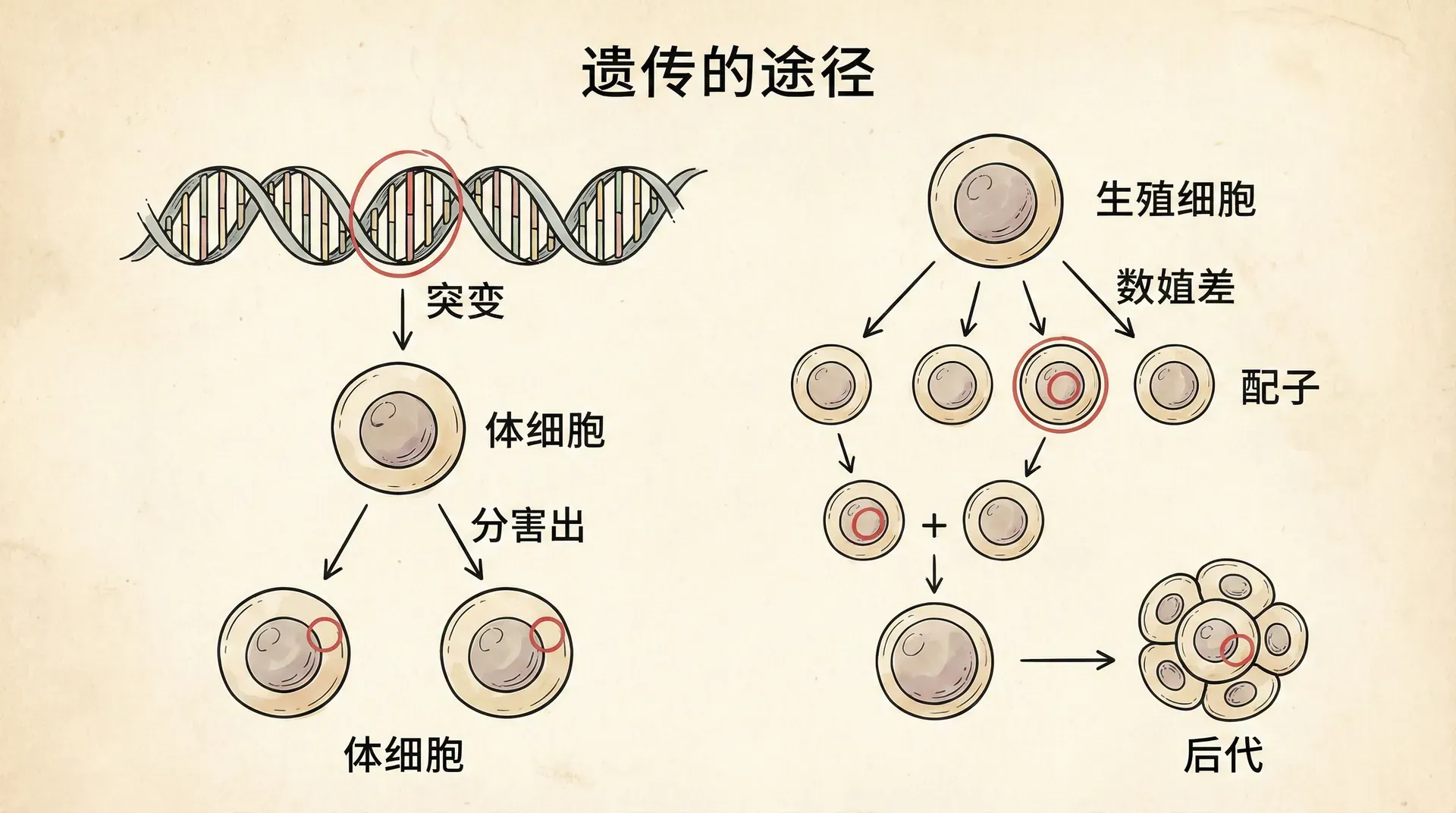
那么,突变是如何在后代中被传递并表现出来的呢?根据现代遗传学的研究,突变往往发生在染色体的某一个特定位点,也就是基因位点。这种突变通常只影响一对同源染色体之一上的一个位置,而不是二者同步改变。也就是说,在一对等位基因中,只有一个发生了改变。
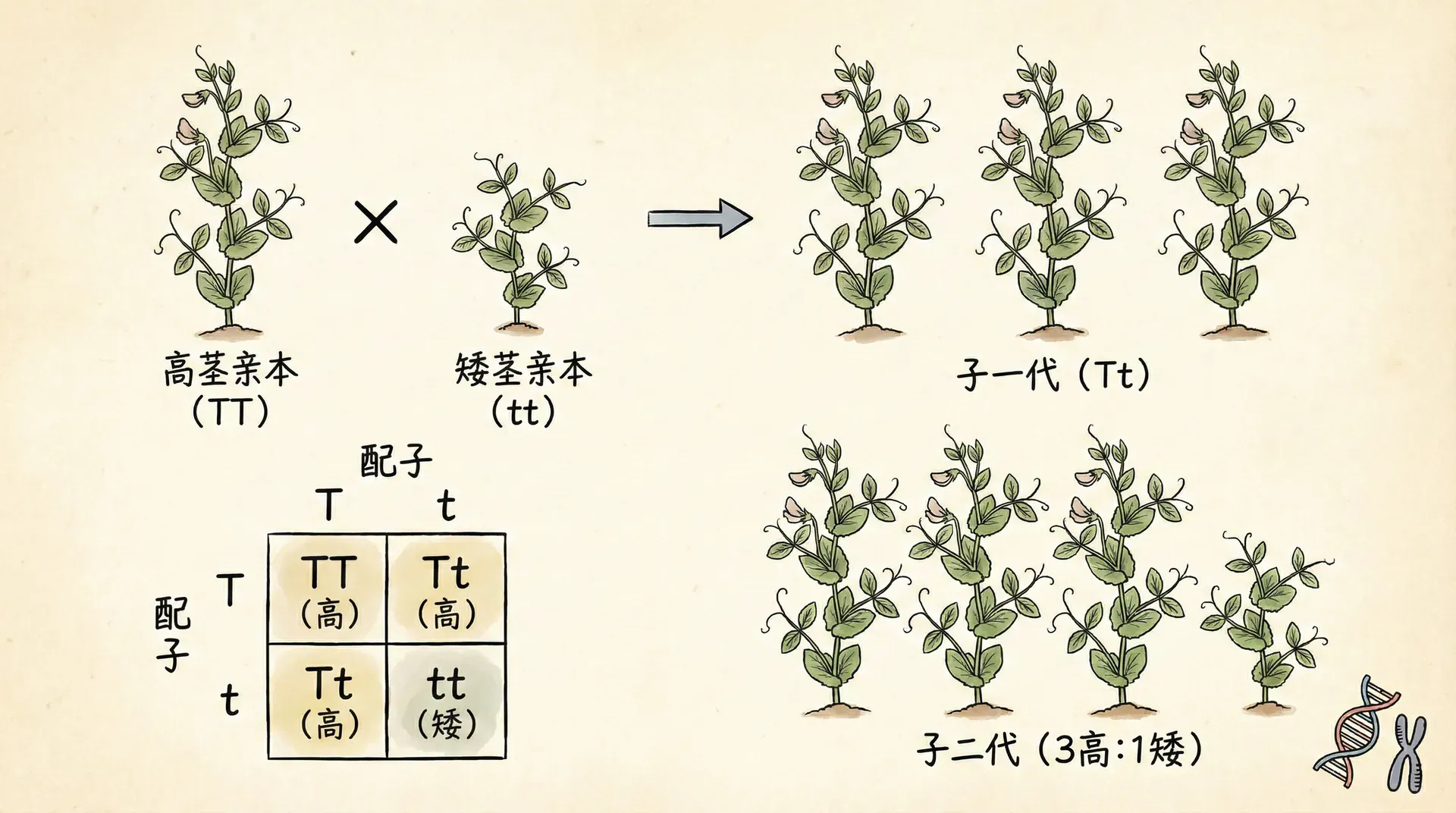
当一个突变体(通常指出现了新性状的个体)与常规个体杂交时,其后代往往出现“性状分离”——有一半后代表现出突变性状,另有一半则维持正常性状。这其实是减数分裂和基因分离规律的直接体现。尤其在F1和F2代,通过观察不同性状的分布比例,可以非常直观地理解遗传的本质。
让我们用图表信息说明:
在实际研究和育种推广中,正是通过反复杂交、选种,不断观察这些性状及其分离比,才逐步揭示了遗传规律的普适性和突变现象的本质意义。
显性与隐性
基本原理
在一次突变事件之后,个体体内存在的“基因脚本副本”就不再相同。按照分子学标准解释,就是同一基因位点上形成了两个不同的序列版本(即两个不同等位基因)。然而,不应简单地将原始序列看作“正统”,突变序列当作“异常”或“异端”——从分子遗传学角度看,这些都是地位完全平等的基因变体。实际上,我们日常看到的“正常”性状本身,在更久远的进化历史中也不过是某次突变的产物。
但在表现层面(即表现型),通常只有一种基因版本会主导个体的外在特征,这一被主导体现出来的版本称为“显性”,而另一个未体现者便称为“隐性”。这个规则并非一成不变,有时还受到环境、修饰基因等影响——但在经典遗传学模型中,这种“显-隐”关系为大多数单基因性状的基本遗传模式。
在自然界中,隐性突变往往比显性突变更为常见,且它们对遗传多样性极其重要。即使在短期内难以直接察觉,它们有可能在后代聚积,当条件合适时,显现出新的遗传和表现型变异。
我们结合中国大型作物改良的案例来分析:比如在玉米品种选育中,若某一植株偶然携带了能够改变籽粒颜色的隐性突变基因,该植株与正常玉米交配时候,杂合的第一代(F1)一般只表现出显性(如黄色)籽粒。但当这些F1植株自由授粉或进行自交,到了第二代(F2),就会有大约1/4的后代表现出“白色”籽粒——这说明只有当两个隐性等位基因在同一植株中聚集(纯合子)时,原先被“隐蔽”起来的隐性性状才得以表现。
这种遗传现象,不仅在玉米广泛存在,在水稻、小麦、豌豆等不同物种中同样出现在性状改良和疾病抗性筛选中,是遗传学实验中的经典规律。
专业术语的介绍
为了让相关讨论更加严谨和规范,我们有必要进一步明确一些基础遗传学的专业术语。前文提到的“密码脚本的不同版本”,在遗传学中被正式称作“等位基因”(allele),即同一基因座上存在的不同序列变体。个体在一个基因座上如果携带两个不同的等位基因(比如一个红花基因,一个白花基因),则称之为“杂合子”(heterozygote);如果携带两个相同的等位基因,则称为“纯合子”(homozygote)。
与此同时,对于个体外在可以观察到的形态或者生理特征,我们称之为“表现型”(phenotype);而决定这些性状的基因组合,则称为“基因型”(genotype)。通过这些术语的规范使用,遗传学家们可以准确描述各种复杂遗传现象。
只有当隐性等位基因同时以纯合状态出现时,其对应的表现型(如白花、白粒等)才能在后代中被观察到。而显性等位基因则无论是处于杂合还是纯合状态都能主导相应的外部性状。这些规律为植物和动物育种、疾病遗传咨询等领域提供了坚实的理论基础和应用手段。
近亲繁殖的危害
隐性有害突变的积累
只要隐性突变还是杂合子状态,它们当然不是自然选择的作用对象。如果它们是有害的(突变经常是有害的),它们也不会被淘汰,因为它们是潜在的。因此,可能积累大量不利突变而不造成直接伤害。但是,它们当然会传递给一半的后代,这对人类、牛、家禽或任何其他我们直接关心其良好身体素质的物种都有重要影响。
假设张先生携带一个隐性有害突变(杂合子状态),因此不会表现出来。假设张太太没有这个突变。那么他们一半的孩子也会携带这个突变——同样是杂合子状态。
只要同样受影响的个体不相互杂交,恶果永远不会显现的危险就不会出现。一旦它们相互杂交,如简单推理所示,它们四分之一的孩子作为纯合子,就会表现出损害。
近亲繁殖的最大危险是兄弟姐妹之间的结婚。他们每个人都有一半的机会潜在地受到影响或不受影响,这些乱伦结合中的四分之一是危险的,因为其四分之一的孩子会表现出损害。因此,乱伦繁殖孩子的危险因子是1:16。
同样地,我这一代两个("纯种")孙子女之间的结合,他们是表兄弟姐妹,其后代的危险因子计算出来是1:64。这些似乎不是压倒性的几率,实际上第二种情况通常是可以容忍的。但不要忘记,我们分析的只是祖辈夫妇("我和我妻子")中一个伴侣的一种可能的潜在伤害的后果。实际上,他们两人都很可能怀有不止一种这类潜在缺陷。
突变是稀有事件的必要性
进化的平衡机制
在谈论遗传学中的变异来源时,我们容易只聚焦于有害突变,毕竟它们似乎带来了更多的健康风险和疾病。但必须正视:突变同样可以是有益的变化。有利的突变能够为生物体提供适应环境的新能力,是生物进化和新物种形成的原材料。
自然界中的自发突变极为罕见。这种稀有性其实起到了平衡的作用。如果突变极其频繁,个体体内同时携带多个新突变的概率就会大幅提高,此时有害突变很容易积累起来,甚至压倒了少数有益突变的正面效应,导致物种整体健康状况下降,甚至面临灭绝的风险。
基因的高度稳定性与持久性是维护物种长期存续的关键,这在工业工程中也有类比。例如在大工厂生产线上,即使要实施技术创新,也需逐项、逐步、更替试验,并确保其它部件保持不变,才能准确评估每项变革的影响。生物体“保守主义”的遗传机制,正是进化选择能有效工作的根本保障。
正因如此,生物体内的基因大部分时间是稳定传承的,只有偶然出现一些微小变异,这些变异才成为自然选择筛选作用的对象。保持突变的稀有性,确保了进化能够逐步推进,既不至于因突变过多而带来灾难,也不会因为完全没有变异而丧失适应环境变化的可能性。这种动态平衡,使得生物进化既有持续性,也有创新性。
X射线诱发的突变研究
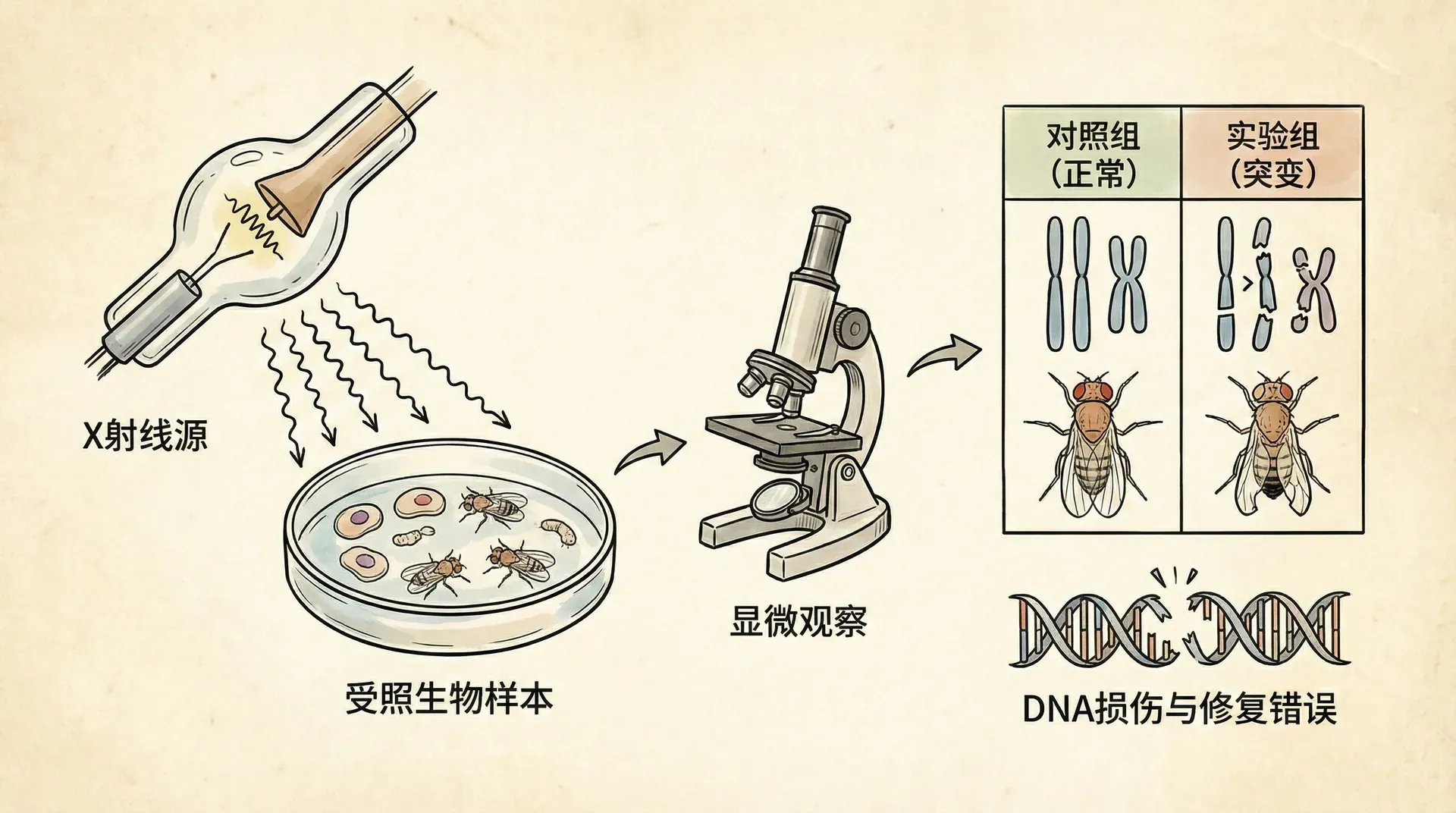
突变率的人工增加
进入20世纪后,科学家尝试人工干预和探究突变过程。在现代遗传学实验中,研究者已经能够通过外来物理或化学因素,例如X射线或γ射线,对生物配子或胚胎进行照射,大幅提升突变发生的概率(即突变率),远超自然界的水平。最著名的研究对象是果蝇(Drosophila),研究者通过大群体培育和追踪,发现使用射线处理可显著增加后代中出现新性状(突变体)的数量。
令人惊讶的是,通过X射线产生的突变,无论在表现、遗传规律上,与自然发生的突变并无本质区别。很多研究发现,相同类型的突变会反复出现,并可被追溯到染色体上特定的区段。正是这些实验,帮助我们理解了突变的分子本质,为基因定位和基因工程打下了基础。
X射线诱发突变的定律
射线诱发突变率的研究揭示了两条简明而深刻的定律:
第一定律:突变数量的增加与照射剂量呈严格的正比,即存在固定的“增加系数”。
这意味着,例如给予一定剂量的X射线后,有千分之一的后代出现突变;而更高剂量下,突变率将线性增长——哪怕是剂量被分成多次单独照射,结果也不会“积累”效应,只与总剂量相关。这一“线性定律”让我们能够以定量方式分析突变风险,揭示突变并非剂量的叠加效应,而是每次照射中染色体受损的单一事件决定。
这一发现推翻了“低剂量多次照射有累加风险”的直观想象,表明突变事件是随机单独事件的结果,那么究竟是什么构成了这一单一事件呢?
第二定律:事件的定位
第二定律:只要照射剂量(以适当的r单位度量)相等,射线种类和能量(波长)差别再大,突变率的增加系数也保持不变。
这意味着,不论是软X射线还是硬γ射线,只要被照射体吸收的总能量(剂量)相同,产生突变的可能性一致。而剂量的标准,一般以创制空气中的电离数为准,因为生物组织和空气在原子成分、反应上相近。
由此推论,真正诱发突变的“单一事件”,是生殖细胞染色体内某个极小“靶标”体积的离子化(或相关分子损伤)。通过分析统计,可以估算出每立方厘米中如果有50,000个离子被创制,某一配子发生特定突变的概率是1/1000,则靶标体积仅占立方厘米的五千万分之一,约等于由10个原子距离构成的立方体,只含大约一千个原子。
这种极微观层次的损伤为后来的分子遗传学埋下了伏笔:后来科学家利用更精细技术,证实确实是DNA分子的特定位点直接损伤或损失,导致遗传突变的发生。
实际应用的考虑
如今,X射线已广泛应用于医学成像、材料检测等诸多领域。人们对直接危害——如皮肤烧伤、辐射致癌、不育等——已有较充分认识与防护,例如手术室、放射科常配备铅屏或防护衣。但在遗传层面,射线仍然可能引起生殖细胞的突变,从而导致下一代出现不显性的遗传病或隐患。
重要的是,即使抵御了射线对个人身体健康的即刻损伤,相关辐射对未来“家族基因库”带来的慢性、隐性累积风险,却极容易被忽视。与近亲繁殖类似,这种隐秘的小概率突变可能在偶然的基因组合下“显现”,威胁种群健康。
例如,如果一位长期暴露于X射线环境工作的护士正好有遗传易感突变,她子孙后代中因为隐性遗传的概率会缓慢增加。虽然这种累积并不需要个人过分担忧,但对于社会整体基因健康,辐射管理和科学普及极为重要。
综合看来,突变不仅奠定了生物多样性和物种进化的理论基石,更在当代医学、遗传优生、防护政策等层面具有现实意义。我们深入理解这些规律,不仅加深了对生命本质的认识,也为人类健康和社会福祉的提升提供了科学参考和行动基础。