种群间相互作用

在自然生态系统中,不同种群之间并非孤立存在,而是通过复杂的相互作用紧密联系在一起。这些种间关系不仅包括直接的相互影响,还通过能量流动、信息传递和生物地球化学循环等多种途径间接地影响着整个生态系统。这些关系塑造了生物群落的结构,调节着物种的分布与多样性,同时也是维持生态系统稳定性和功能多样性的基础。
种群间的相互作用形式多种多样,包括但不限于竞争、捕食、捕食者-猎物、寄生、互利共生、偏利共生和拮抗等。在现实生态系统中,这些关系往往并非单一存在,而是交织成一个复杂的网络。例如,某些鸟类既会与同类竞争食物资源,又可能作为捕食者以昆虫为食,还会为树木传播种子与其形成互利共生关系。正是这些错综复杂的相互作用,让生态系统具备了高度的动态平衡与适应性。
此外,随着全球化进程的加快和人类活动的加剧,生物入侵问题日益严峻。外来物种入侵往往会打破原有的生态平衡,造成本土物种的数量锐减甚至灭绝,影响原有生态系统的稳定性和多样性。最后,本章将结合中国实际案例,重点讨论生物入侵对本土物种的威胁,解析入侵物种如何通过竞争、捕食、寄生等多种机制影响原有种群,并简要介绍防控措施与管理策略,以期为生态保护提供科学支持。
竞争关系与生态位理论
当多个种群利用相同的有限资源时,就会产生竞争关系。竞争是生态系统中最常见的种间关系,对种群数量、分布及生态系统结构和功能产生深远影响,也促使物种进化和生态位分化。
竞争的类型与机制
种间竞争通常分为两类:资源竞争(间接)和干扰竞争(直接)。下面总结两种主要竞争类型及相关实例:
在中国东北的玉米田生态系统中,玉米与苘麻杂草竞争土壤水分和氮素。当苘麻密度升高至每平方米25株时,玉米产量显著下降。因此,田间管理中常需定期除草,以确保作物竞争优势。
竞争强度受资源稀缺程度及物种生态位重叠度影响。当资源充足时竞争较弱,资源不足则形成激烈竞争,甚至出现竞争排斥。
生态位的概念与分化
生态位是物种在生态系统中的角色与利用的综合环境条件。包括基础生态位(无竞争时能利用的空间和资源)和实际生态位(竞争存在下实际利用的空间和资源)。
高斯竞争排斥原理认为,生态位完全重合的物种无法长期共存。为减少竞争,自然界中许多共存物种形成“生态位分化”,即在资源利用、栖息地或活动时间等方面产生差异。
以云南橡树林为例:亚洲象和野猪虽然在相同林区活动,但亚洲象以竹笋、树叶为主食,偏好湿润河谷,而野猪取食地下块茎、果实,活动空间偏向林缘和耕地。这种食性和空间分化,促成两物种在资源有限时长期共存。
竞争共存的机制
尽管理论上生态位高度重叠物种难以共存,但自然界中共存现象广泛,说明存在促进共存的机制。主要包括资源分割、时间分割和空间分割等。
- 资源分割:不同物种利用不同类型、大小或空间的资源。例如,长白山针叶林中,红松主分布于土壤肥厚区域,云杉偏向阴湿区,落叶松在较干旱地带优势明显。
- 时间分割:物种在不同时间段利用同一资源。如下表所示:
图中可以看到,不同植物群组通过在不同季节达到生长高峰,实现了对资源的错开利用,从而减少彼此的直接竞争,提升了群落的整体生产力。
捕食与被捕食的动态关系

捕食关系是指捕食者以其他物种为食,是生态系统能量流动与结构形成的核心机制。捕食不仅影响被捕食者种群,还促进捕食者和被捕食者间的复杂适应性协同进化。
捕食者与被捕食者的数量动态
捕食者和被捕食者在种群数量上表现为典型的周期性波动。当被捕食者数量增加时,捕食者数量随之增长,而过多的捕食又减少被捕食者种群,进而影响自身数量。
以加拿大猞猁和雪兔为例,雪兔种群大约每10年达到一次高峰,猞猁种群的波峰落后雪兔1-2年。类似的动态关系在我国新疆荒漠—草原区的狐狸与野鼠、祁连山的雪豹与岩羊之间也可观测到。
上图展现了捕食系统中捕食者与被捕食者数量的典型波动特征,二者都受到相互影响,表现出时滞性。
被捕食者的防御策略
被捕食者进化出多样化的防御手段,包括形态、化学和行为等方面,常用下列表格总结:
拟态是一类特殊防御,包括贝氏拟态(无毒种模仿有毒种)和缪勒拟态(多有毒种彼此模仿,协同强化警戒效果)。
如云南西双版纳的枯叶蝶,其翅膀外观极似干枯树叶,能有效欺骗捕食性鸟类。贝氏拟态的例子如亚洲眼蝶无毒却模仿有毒蝶类体色,从而减少被捕食风险。
捕食者的捕猎策略
捕食者为提高捕食效率,演化出多样猎食策略,列举如下:
在中国山东沿海,灰喜鹊曾被观察到集体围捕灰鼠,采用合作策略成功获取比单独行动时更大的猎物。
通过这些多样的捕食与防御策略,捕食者与被捕食者之间不断进化与适应,推动了生态系统结构与多样性的形成。
寄生与共生的生态学意义
除了竞争和捕食,生物之间还有更加微妙和复杂的关系,比如寄生、各种类型的共生等。这些联系是生态系统多样性和稳定性的重要基础。
寄生关系
寄生是指一种生物(寄生者)生活在另一种生物(宿主)体表或体内,依赖宿主摄取营养、对宿主产生伤害但通常不会立刻致死。寄生者往往体型较小、繁殖较快,而宿主体型较大。
下方是寄生的主要类型与典型举例:
寄生关系广泛存在于自然界。比如中国东北的水牛经常被蜱虫寄生,不仅吸血,还可能传播病原体。又如蛔虫在人体肠道内寄生,影响人体健康。
寄生者不仅调控宿主种群数量,还会改变种间关系格局。例如,当两个物种竞争时,若其中一个受寄生压力更大,就可能在竞争中处于劣势,“竞争排斥”由寄生关系改变。
例如,冬虫夏草(虫草菌)专门寄生在蝙蝠蛾幼虫体内,最终杀死宿主并从其头部长出子实体。这展示了寄生对宿主具有终极影响,但寄生者必须在宿主死亡前完成营养积累和繁殖。
共生类型与实例
共生指两种生物长期生活在一起、相互作用的现象。按双方受益程度不同,常见类型如下:
共生关系推动生物进化许多复杂功能,比如动物消化道里的微生物能分解纤维素、合成维生素、抑制病原。人类肠道的细菌总数万亿级,总重约1–2公斤,对健康至关重要。
清洁共生与传粉关系
清洁共生是一种典型的互利共生。如下:
在珊瑚礁区,清洁鱼开设“清洁站”,连凶猛的石斑鱼也安静地接受服务。在中国南方,牛背鹭常见于水牛背上捉虫,成为田野独特景观。
传粉互利关系则极为关键,直接关系到植物的有性繁殖。以榕树与榕小蜂为例:
- 榕树为榕小蜂提供产卵和生育空间、食物(花序)
- 榕小蜂专为榕树授粉,完成传粉和繁殖使命
每种榕树几乎都配有唯一的榕小蜂种类,互相依赖,缺一不可。这种“专一共生”在中国南方尤为丰富,展现了植物与传粉昆虫间极其精密的协作。
种间关系的定量模型
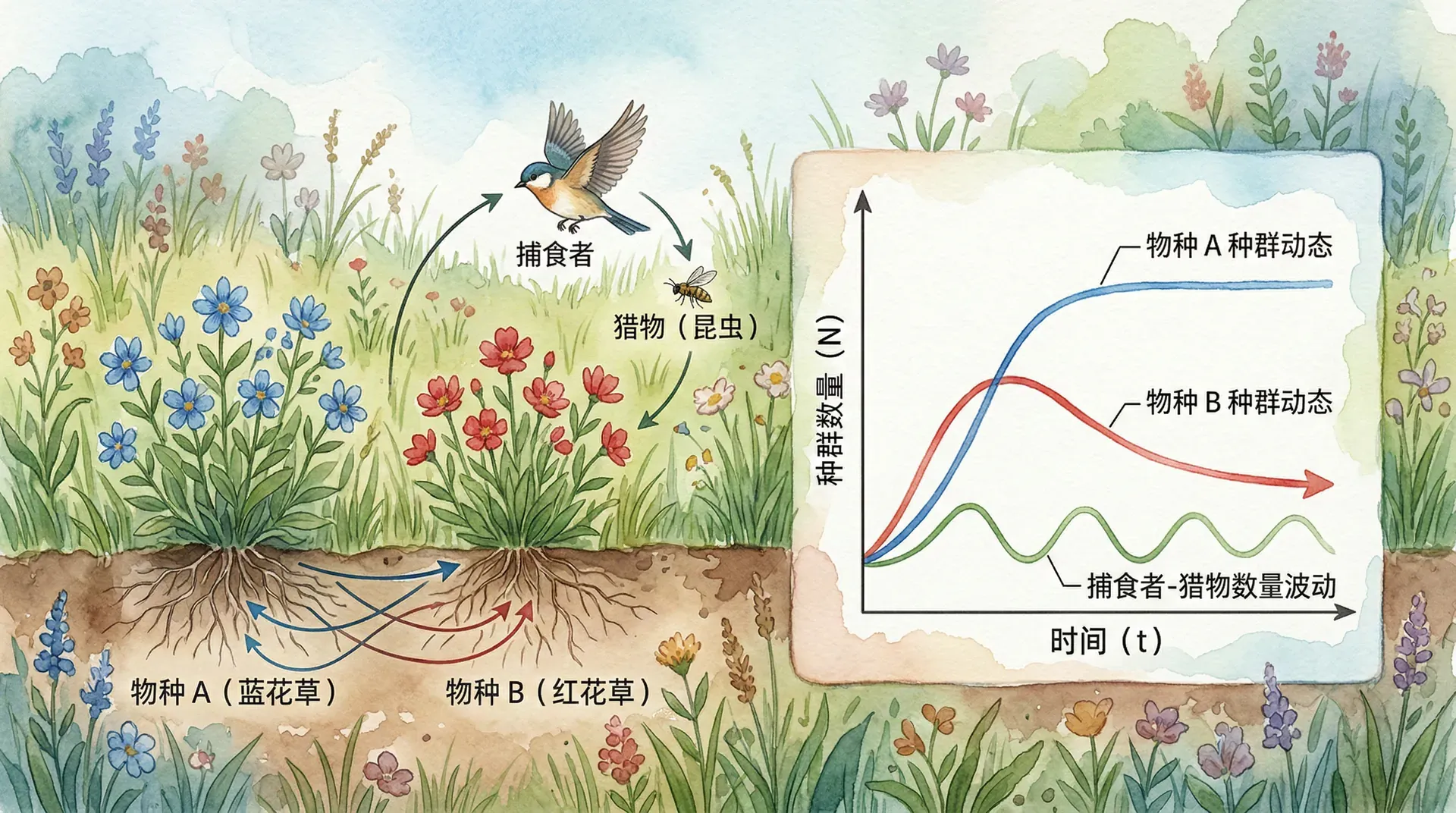
为了深入理解种间相互作用的动态过程并进行预测,生态学家建立了多种数学模型用于定量描述这些关系。虽然这些模型对自然规律进行了简化,但能够揭示种间关系的核心机制,并为生态管理实践提供理论支持。
竞争模型
洛特卡(Lotka)和沃尔特拉(Volterra)在20世纪20年代分别提出了描述两个物种间竞争关系的经典模型。它是在逻辑斯谛增长模型的基础上,增加了种间竞争的抑制项。对于两个竞争物种,数学表达如下:
其中,、分别为物种1和物种2的种群数量,、为内禀增长率,、为环境容纳量,和为竞争系数,反映物种间的竞争强度。
竞争结果依赖于和的取值,部分预测结果可用下表总结:
举例:在池塘环境里,水葫芦与睡莲两种植物争夺阳光和空间,若水葫芦竞争能力显著强于睡莲,则模型可成功预测水葫芦会占据优势,甚至将睡莲排斥。
这些模型为“竞争排斥原理”和生态位分化提供了理论基础。
捕食模型
洛特卡-沃尔特拉捕食-被捕食模型刻画了捕食者与被捕食者相互作用的动态。其核心数学表达:
其中,为被捕食者数,为捕食者数,为被捕食者内禀增长率,为单位捕食率,为转化效率,为捕食者死亡率。公式中的表示捕食作用导致的被捕食者减少以及捕食者生物量增加。
主要参数意义如下:
此模型预测捕食者与被捕食者数量会出现周期性波动。例如:草原上的狼和兔子,兔子数量因缺乏天敌而上升→狼数量增多→兔子被捕食减少→狼随之减少,随后兔子恢复,如此循环。
为更贴近现实,科学家引入了“功能反应”,即捕食者单位时间内的捕食速率如何随被捕食者密度变化,其典型类型如下:
例如,瓢虫捕食蚜虫时,刚开始蚜虫很少时不易发现,蚜虫密度升高时捕食速率逐步提升,属于III型反应。
模型在实际中的应用
这些定量模型在自然资源管理与保护中具有重要指导价值。例如:
- 渔业管理:利用种群动态模型预测鲢鱼等捕捞物种最佳产量,东海带鱼捕捞配额即基于此类模型,防止资源衰退。
- 害虫防治:棉田里利用捕食-被捕食模型分析瓢虫与蚜虫关系,预测放养瓢虫所需阈值,使蚜虫常年处于无害水平,减少农药。
- 保护生物多样性:预测新引入天敌是否可稳定抑制入侵物种,如北美引入天牛控制紫茎泽兰。
数学模型建立在一定的假设之上(如环境均一性、参数不变等),实际应用中需结合实地调查和实验数据进行调整。生态系统随机扰动、气候变动、多物种网络等因素都可能导致预期与实际结果差异。因此,模型应作为管理和决策的有力工具,但不能作为唯一依据。
生物入侵与本土物种保护
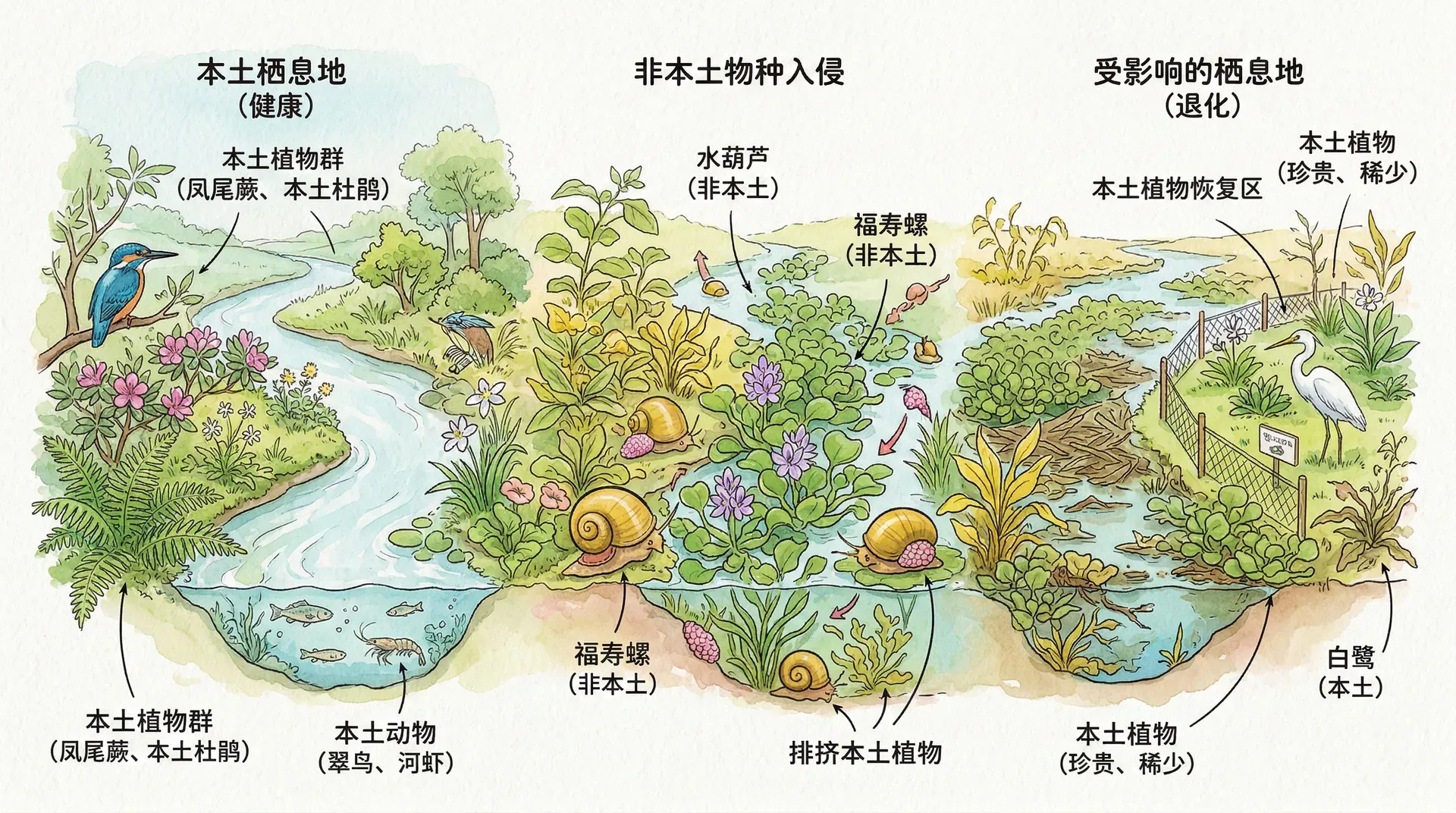
随着全球化和人类活动的加剧,生物入侵已成为威胁生物多样性和生态系统稳定的重大因素。外来物种由于缺乏天敌,常常能迅速扩散,给本土生物和生态系统带来严重影响。生物入侵大致可分为引入、定居、扩散和影响四个阶段:物种首先通过有意(如作物、家养动物)或无意(如随货物、船舶等)途径进入新环境,能够适应并存活下来的物种形成种群,逐步向更广区域扩展,最终产生生态影响,包括资源竞争、传播疾病、改变栖息地等。
中国受生物入侵影响尤为严重,已记录外来入侵物种600多种,其中上百种对生态与农业危害巨大,涵盖植物、动物、微生物等类群,分布广泛。下面是几种典型案例:
- 水葫芦:原产南美,20世纪初引入中国,因强大繁殖力在南方水域泛滥,影响水生态、航运与灌溉,治理成本高。
- 松材线虫:北美原产,通过寄主天牛传播,能在短时间致松林大面积死亡,自1980年代以来扩散全国,危害森林及风景区。
- 美国白蛾:1979年首次发现于辽宁,危害农林、城市绿化,繁殖与扩散能力强,叶片啃食严重甚至导致整树死亡。
- 福寿螺:南美原产,起初为食用引入,后因弃养在南方水田泛滥,危害水稻及水生态。
- 紫茎泽兰:1930年代传入云南、贵州等地,具化感作用,迅速扩展,导致草地退化、牧草减少,影响畜牧业。
生物入侵不仅抑制本土物种、甚至引发灭绝,还会改变环境结构与功能。例如紫茎泽兰凭借化感作用与高繁殖力,压制本土草本植物,导致牧草锐减;加拿大一枝黄花能使群落单一化,削弱动物栖息和食物资源;桉树因生长快、耗水多且落叶分解慢,被大量种植后会耗竭土壤水肥,改变水分与养分循环,造成“抽水机”“抽肥机”问题。
保护本土物种必须采取综合策略:
公众的参与至关重要。增强对生物入侵危害的科普,倡导不随意丢弃外来宠物或植物,及时报告新发现物种等,将极大提升防控成效。入侵物种的扩散很多与人的不当行为有关,只有全社会协作,才能更好地遏制生物入侵。
总结
生态系统中,竞争、捕食、寄生和共生等多种关系协同作用,塑造了生物群落的结构,维持着生态系统的动态平衡。例如,种间竞争能够决定物种的分布与多样性,捕食关系调节着种群数量,寄生和共生则展现了物种间互为影响和依赖的生态网络。这些种间关系不仅发生于动植物之间,更广泛涉及微生物与高等生物的相互作用。
在分析这些复杂关系时,使用数学模型能帮助我们定量描述物种间的相互影响、动态变化以及影响生态系统结构的关键因子。模型如洛特卡-沃尔泰拉模型、功能反应模型等,让我们能够预测生态系统在不同条件下的可能变化趋势,同时也为管理与保护提供科学依据。
当前,生物入侵已成为全球范围内生态安全的重要威胁。外来物种通过竞争、捕食、寄生等方式严重威胁本土生物和生态平衡,甚至可能导致本地物种灭绝和生态系统功能损失。面对这一挑战,我们应采取综合性的防控措施,包括加强检疫、早期监测和快速响应、开展科学的防控和生态恢复。公众参与同样不可忽视,通过加强宣传教育、提升民众环保意识,可以有效遏制生物入侵的蔓延。
深入理解种间关系不仅有助于我们认识自然界的运行规律,还为生物多样性保护、农业生产可持续发展以及自然资源科学管理提供了坚实的理论基础和实践指导。今后,我们还需不断深化对生态系统中种间关系的研究,结合实际问题,探索更有效的生态保护与管理路径,从而实现人与自然和谐共生的目标。
本节练习
1. 某森林中有A、B两种鸟类,它们都以昆虫为食。A鸟主要在树冠层活动,捕食树叶上的昆虫;B鸟主要在树干活动,捕食树皮裂缝中的昆虫。这两种鸟能够在同一片森林中共存的主要原因是什么?请从生态位理论的角度分析。
答案:A、B两种鸟类能够在同一片森林中共存,主要是因为它们发生了生态位分化。虽然两种鸟都以昆虫为食,但它们在觅食空间(A在树冠层,B在树干)和食物类型(A捕食树叶上的昆虫,B捕食树皮裂缝中的昆虫)上存在差异。这种生态位分化减少了两个物种之间的资源竞争,使它们能够共享同一片森林资源而不发生激烈竞争。这体现了竞争排斥原理的作用:当两个物种生态位完全重叠时会产生激烈竞争,而通过生态位分化,物种可以减少竞争强度,实现共存。这种分化可能是长期进化的结果,也可能是当前环境条件下的行为调整。
2. 在某草原生态系统中,草是主要生产者,野兔以草为食,狼以野兔为食。假设某年由于干旱导致草的生长量大幅下降。请预测这一变化对野兔和狼种群数量的影响,并解释原因。
答案:草生长量下降会导致野兔种群数量减少,进而引起狼种群数量下降。具体影响过程如下:
首先,草生长量下降直接减少了野兔的食物供应,野兔因食物不足,营养状况变差,繁殖率下降,死亡率上升,导致野兔种群数量减少。
随后,野兔数量的减少使得狼的食物来源减少,狼会面临食物短缺的问题。捕食压力增大但猎物减少,狼的捕食成功率下降,部分狼可能因饥饿而死亡,或因营养不良导致繁殖率下降,从而使狼种群数量也随之减少。
这个过程体现了生态系统中的营养级联效应和自下而上的控制机制。生产者(草)的变化通过食物链向上传递,影响初级消费者(野兔)和次级消费者(狼)。这也说明了生态系统各组分之间的相互依赖关系,底层营养级的变化会对整个食物链产生连锁反应。
3. 豆科植物与根瘤菌的关系属于哪种种间关系?这种关系对农业生产有什么实际意义?
答案:豆科植物与根瘤菌的关系属于互利共生关系。
在这种关系中,根瘤菌侵入豆科植物根部形成根瘤,在根瘤中将空气中的氮气(N₂)转化为植物可以吸收利用的氨态氮(NH₄⁺),为植物提供氮素营养。作为回报,豆科植物通过光合作用产生的糖类等有机物为根瘤菌提供能量和生长所需的碳源。双方都从这种关系中获益。
这种关系对农业生产的实际意义包括:
-
豆科植物可以自行固氮,减少对氮肥的依赖,降低农业生产成本和环境污染。氮肥的生产和过量使用会造成能源消耗和环境问题,而生物固氮是环保且经济的氮源。
-
豆科作物(如大豆、花生、苜蓿等)与禾本科作物(如水稻、小麦、玉米等)进行轮作,可以通过根瘤菌固氮提高土壤肥力,改善土壤结构,为后茬作物提供氮素,实现可持续农业生产。
-
豆科绿肥植物(如苜蓿、豌豆等)可以种植后翻压入土,作为天然有机肥料,增加土壤有机质和氮含量,这种方法在有机农业中广泛应用。
4. 下表是某地区引入外来物种后本土植物种类数量的变化数据。请分析该外来物种是否成为入侵物种,并说明判断依据。
答案:根据表中数据,该外来物种已经成为入侵物种。判断依据如下:
-
外来物种的覆盖率持续快速上升,从2010年的5%增长到2018年的58%,8年间增长了11倍多,显示出极强的扩散能力和竞争优势。
-
随着外来物种覆盖率的增加,本土植物种类数持续减少,从2010年的85种降至2018年的58种,减少了约32%。这表明外来物种的扩张对本土植物造成了严重的负面影响。
-
外来物种覆盖率与本土植物种类数呈现明显的负相关关系,说明外来物种的扩张直接导致了本土植物多样性的丧失。外来物种可能通过资源竞争、化感作用或改变生境条件等方式排挤本土植物。
-
这种快速扩散和对本土生态系统的显著负面影响符合入侵物种的典型特征。入侵物种通常具有生长快、繁殖能力强、适应性广等特点,能在短时间内占据大量生态空间,威胁本土生物多样性。
因此,需要对该外来物种采取紧急控制措施,防止其进一步扩散,并通过生态恢复措施保护和恢复本土植物群落。
5. 在某水域进行鱼类养殖时,如果单养一种鱼类,产量为500kg/亩;如果混养草鱼、鲢鱼和鳙鱼三种鱼类,总产量可达900kg/亩。请从生态位理论的角度解释混养产量更高的原因。
答案:混养产量更高的原因在于三种鱼类发生了生态位分化,更充分地利用了水域中的资源,提高了整体资源利用效率。
草鱼主要以水草、陆生植物和人工投喂的饲料为食,属于草食性鱼类,主要在水体中下层活动。鲢鱼以浮游植物(如硅藻、绿藻等)为主要食物,通过滤食方式摄食,主要在水体上层活动。鳙鱼(又称胖头鱼)主要以浮游动物(如轮虫、枝角类等)为食,也采用滤食方式,活动范围包括中上层水域。
这三种鱼类在食物来源(水草、浮游植物、浮游动物)、摄食方式(啃食、滤食)和活动水层(上层、中层、下层)上存在明显差异,它们的生态位重叠度低,种间竞争较弱。混养时,每种鱼类都能充分利用自己擅长利用的资源,而不会对其他鱼类的资源造成过多竞争。
相比之下,单养一种鱼类时,只能利用水域中的部分资源,其他资源未被充分利用,造成资源浪费。混养通过生态位互补,实现了对水域资源的立体化、多层次利用,因此总产量更高。
这个例子体现了生态位理论在实际生产中的应用。通过合理搭配不同生态位的物种,可以提高生态系统的生产力和资源利用效率,这一原理不仅适用于渔业,也广泛应用于农业、林业等领域。
6. 捕食者和被捕食者之间存在“军备竞赛”式的协同进化。请举例说明这种协同进化如何推动双方的适应性进化,并讨论这对维持生态系统平衡有何意义。
答案:捕食者与被捕食者之间的协同进化是指双方在长期相互作用中,一方的适应性改变促使另一方发展相应的对抗策略,形成持续的进化“军备竞赛”。这种协同进化推动了双方的适应性进化,并对维持生态系统平衡具有重要意义。
协同进化的典型例子包括:猎豹与羚羊的速度竞赛。猎豹进化出了极快的奔跑速度(可达110公里/小时)来追捕猎物,而羚羊也进化出了高速奔跑和灵活转向的能力来逃避捕食。那些跑得更快的猎豹有更高的捕食成功率和生存繁殖机会,而跑得更快的羚羊有更高的逃生概率,结果是双方的速度都在进化压力下不断提升。
在中国的例子中,蝙蝠与蛾类之间的超声波"军备竞赛"令人惊叹。蝙蝠进化出了回声定位系统,通过发射超声波并接收回声来探测和追踪飞行中的昆虫。作为应对,许多蛾类进化出了能够听到蝙蝠超声波的听觉器官,一旦探测到蝙蝠的超声波信号,就会采取紧急回避动作如急速下降、不规则飞行等。更有甚者,某些蛾类还进化出了干扰蝙蝠回声定位的能力,能够发射自己的超声波信号混淆蝙蝠的探测。作为反击,蝙蝠又进化出了更复杂的超声波调制方式和更精密的信号处理能力。这种相互促进的进化推动了双方感觉和神经系统的高度发展。
这种协同进化对维持生态系统平衡具有重要意义:
-
协同进化防止了一方完全压倒另一方。如果捕食者的捕食能力无限增强而被捕食者没有相应的防御进化,被捕食者将被捕食殆尽,随后捕食者也会因失去食物而灭绝。协同进化使双方保持相对平衡的竞争关系,两者都能长期共存。
-
协同进化促进了生物多样性。在进化压力下,物种会分化出多样的捕食策略和防御策略,增加了生态系统的复杂性和物种多样性。不同的捕食者专门化于捕食不同的猎物,不同的被捕食者发展出不同的防御机制,形成了复杂的食物网结构。
-
协同进化提高了生态系统的稳定性。多样化的种间关系和复杂的食物网使得生态系统更具韧性,能够更好地应对环境变化和扰动。如果某一物种数量波动,复杂的种间关系网络可以缓冲这种波动对整个系统的影响。
因此,捕食者与被捕食者的协同进化不仅是生物进化的重要机制,也是维持生态系统动态平衡和生物多样性的关键过程。理解这一过程有助于我们更好地认识自然界的复杂性,为生态保护和管理提供理论指导。