基因的远距离控制力
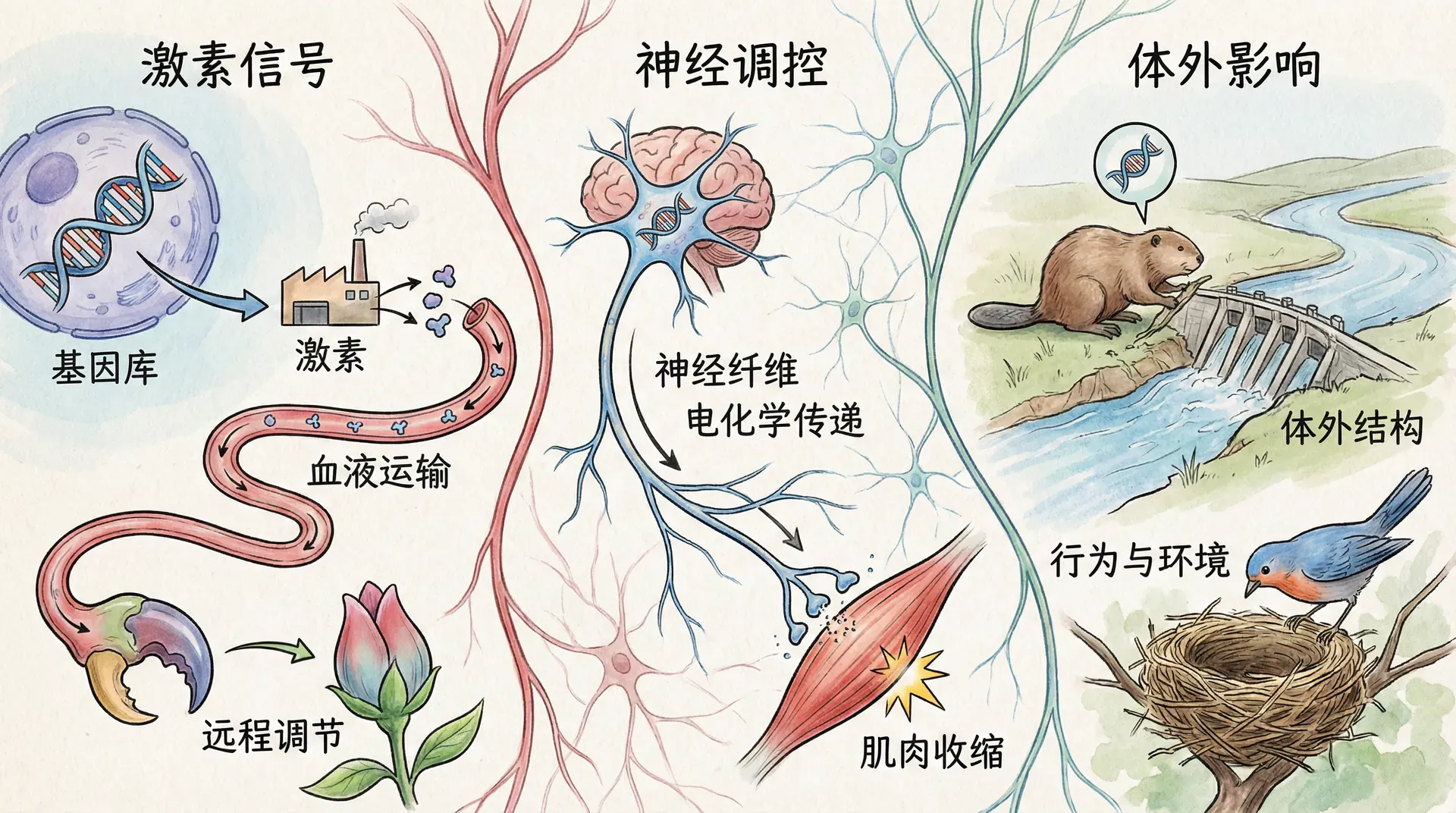
生命科学的重大转变之一,是我们逐渐认识到:基因的作用已远远超越了单个生物体自身。过去,我们惯常认为基因只负责决定个体的形态和性状,但现代研究表明,基因的影响实际上会通过复杂的机制延展到外部环境与其它生物,塑造更广阔的生态格局。
比如一只蝴蝶的飞舞,不仅展现了基因塑造的美丽外观,更体现了进化赋予它与花朵互动、参与授粉网络的能力。这些基因影响还可以进一步扩展,影响花期、传粉行为,乃至生态系统资源的分配和能量流动。类似地,虫草菌可精准控制昆虫的行为完成生命周期轮换,寄生虫能在不同宿主间实现遗传策略的传播;大熊猫的食性和行为影响了竹林生态结构,进而反馈到整个保护区的管理和植物遗传多样性;蜜蜂基因促成蜂巢的构建,又塑造了小范围内的微型生态系统。
这些案例表明,所谓“延伸表现型”理论,强调的是基因通过影响行为、结构乃至环境,形成跨越个体、种群甚至生态系统的连锁反应。理解这一观点,有助于我们跳出生物与环境、个体与群体彼此割裂的旧有框架,更深入把握进化和生态系统动态背后的复杂动力与协作机制。
基因与个体的关系
传统观念的挑战
传统生物学多认为,基因仅作为个体生命的组成部分,把它们的作用限定在生物体自身,仿佛一个工厂的影响只能局限在厂房内部,影响不到更广阔的外部世界。基因只影响眼睛的颜色、体型、毛发等自身性状,这种认知在很长时间内主导了我们的思考方式。
但现代研究却逐渐推翻了这种“封闭模型”。我们发现自然界处处都是基因“远距离操作”的实例。基因的表达和产品并不是以身体为终点,而是能够溢出边界,在群体、种群甚至生态系统层面产生深刻的连锁反应。
基因的影响力就像无线电波一样,可以穿越个体的边界,在更广阔的范围内发挥作用。
以中国著名的旗舰保护物种大熊猫为例。大熊猫的基因本应只作用于自身(如决定其黑白相间的毛色、健壮的体魄、能够消化竹子的特殊消化酶系统等),但如果我们观察得再深一层,不难发现它们的基因实际上还间接塑造着周边的竹林生态。它们对特定竹种的偏好推动了某些竹种的繁茂生长,影响了竹林的空间格局,有些竹种为适应大熊猫的觅食压力也发生了遗传改变。这种基因-环境-生态圈层的相互作用正是延伸表现型的活生生例证。
甚至连人类社会也被间接影响:正因大熊猫的独特生态位和保护价值,我们投入了大量资源植被修复,推动了生态旅游和科学研究,从而影响了保护区范畴、当地经济以及公众的环保意识。这些外溢影响层层递进,打破了基因“只属于自身”的传统思维。
表现型概念的拓展
生物学中,“表现型”传统上被定义为由基因控制、在个体上观察到的特征,比如眼睛的颜色、身高、免疫能力或典型行为。这种认识类似于只关注一颗种子长成植株,而忽略了它如何通过根系、分泌物、遮荫效应影响周边土壤微生物、邻近植物甚至昆虫群落。事实上,基因的作用远远不止于可见特征,而是基于复杂网络的多层次协同,能够深远改变环境结构和生态交互。
真正的基因影响力更像是由个体出发,逐渐外溢到近邻、群体、环境和更大生态系统的连续梯度。在生物体内部,基因的控制力最强;但随着物理距离和生物层级的提升,这种控制力虽渐行渐弱,却并不会完全消失,反而以多样化和间接化的方式持续发挥作用。例如,蜜蜂建巢的天性、白蚁社会的分工、鸟巢构造都是基因间接影响外部世界的方式。
这种对于表现型的拓展理解,让我们必须重新思考“生物体边界”以及“遗传效应止于何处”的传统观念。它不仅使我们能够解释如寄生虫行为操控、共生体形成等生物学奇观,也为现代生物工程、保护生态学等领域带来了重要启示。
超越边界的基因影响
基因的建筑才能
基因不仅塑造了生物体本身,还能延伸出超出身体界限的结构和功能。蜜蜂建造蜂巢的行为受到基因控制,而蜂巢的形状、材质选择和空间布局,都是基因延伸表现型的体现。蜂巢不是蜜蜂身体的一部分,但它同样是基因表达的产物。这种认识让我们意识到,基因的“产品”不仅包括生物体本身,还包括生物体创造的各种结构。
另外,在中国南方常见的织布鸟(黄胸织布鸟)也展示了基因的建筑才能。雄鸟编织的精美鸟巢,其复杂程度不亚于人类的手工艺品。编织技巧、材料选择、悬挂方式,这些看似学习得来的技能,实际上深深烙印在基因中。
跨物种的基因控制
更令人惊讶的是,基因的影响可以跨越物种边界。中国常见的菟丝子(Cuscuta chinensis)为我们提供了一个典型例子。这种寄生植物的基因不仅控制自身的生长,还能影响宿主植物的生理机能。
菟丝子通过分泌特殊化学物质,改变宿主植物的新陈代谢,使宿主为自己提供更多营养。从某种意义上说,菟丝子的基因正在“遥控”宿主植物的身体机能。
因此,基因的影响力可以跨越物种边界,一个生物体的基因能够控制另一个生物体的生理状态和行为。
寄生与共生
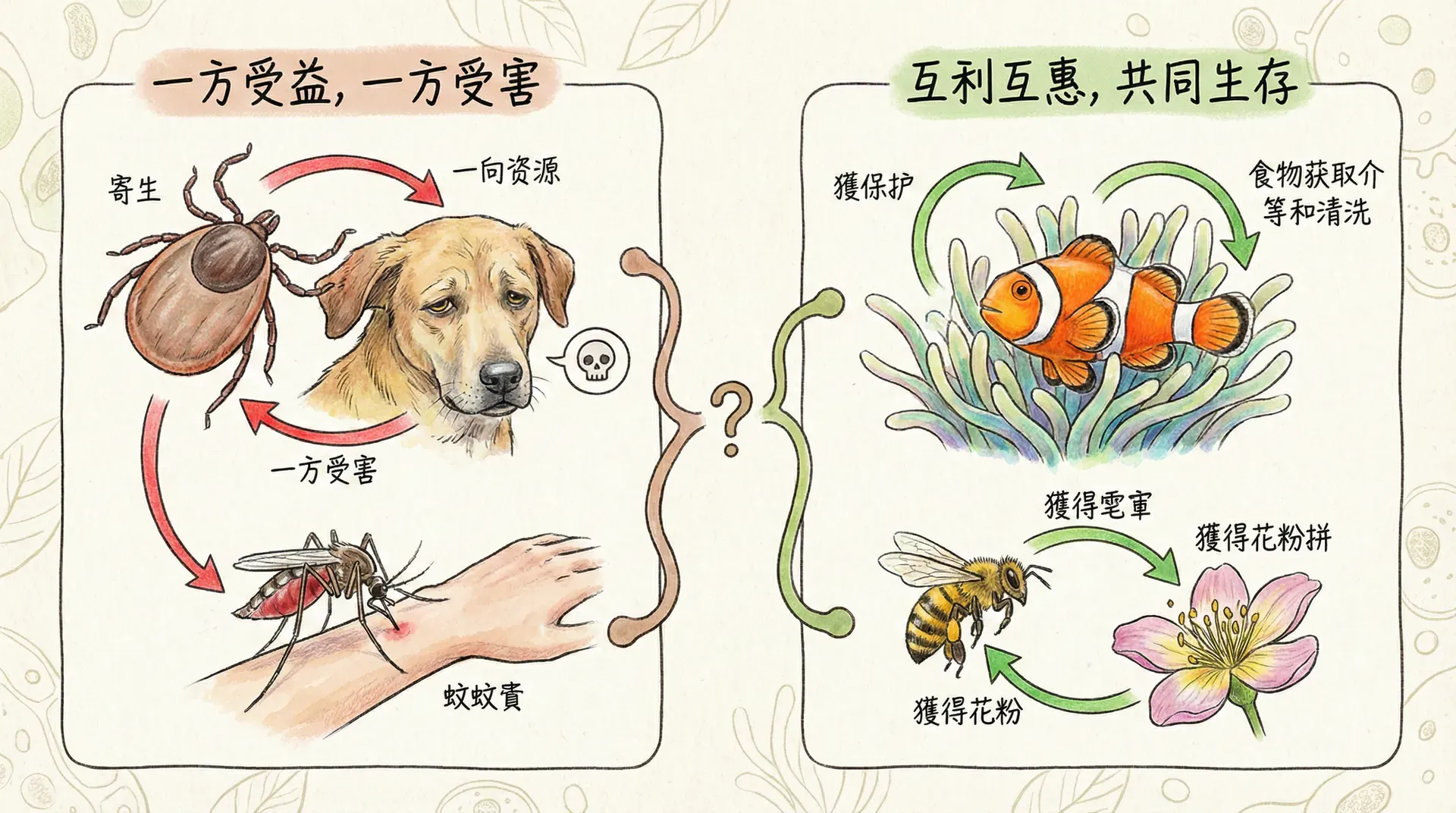
利益一致时的合作
当不同物种的基因拥有共同的传播途径时,它们会表现出惊人的合作性。以中国常见的地衣为例,这实际上是真菌和藻类的共生体。真菌的基因和藻类的基因在进化过程中达成了“协议”——共同创造一个适合双方生存的环境。
真菌提供结构支撑和水分保持,藻类进行光合作用提供养分。两套完全不同的基因系统协调工作,共同塑造了地衣独特的形态和生理特征。
冲突中的操控
在自然界中,不同物种的基因利益往往并不一致,当它们发生冲突时,会出现复杂的“博弈”与“操控”现象,尤以寄生与宿主的关系为最典型。在中国,常见的例子有植物线虫对植物根系的操纵、真菌操控昆虫行为,以及寄生蜂影响瓢虫行为等。下面简明总结几类典型的冲突操控案例及其机制:
以奥菲奥科迪斯菌感染蚂蚁为例:真菌孢子可精准操控蚂蚁的大脑和神经系统,使其脱离蚁群、攀爬植物高处,在叶片背面咬住固定并死亡。死亡蚂蚁的身体成为孢子释放平台,帮助真菌广泛传播。可以说,真菌的基因充当了操控“遥控器”,主导宿主体的行为为自己利益服务。
这些操控现象同样见于病毒、细菌和各种动物。例如,寄生蜂让瓢虫变身“守护者”,或某些寄生虫通过气味改变吸引新的宿主。它们共同说明:生物体不仅仅受自身基因支配,还是不同复制子在利益冲突下的“博弈场”。
这些跨物种的操控案例提醒我们,生物体的行为有时是“外来基因”的产物,而非仅仅服务自身利益。生态系统中许多奇特行为,其实是不同基因间暗中较量和妥协的结果。
复制子与载体
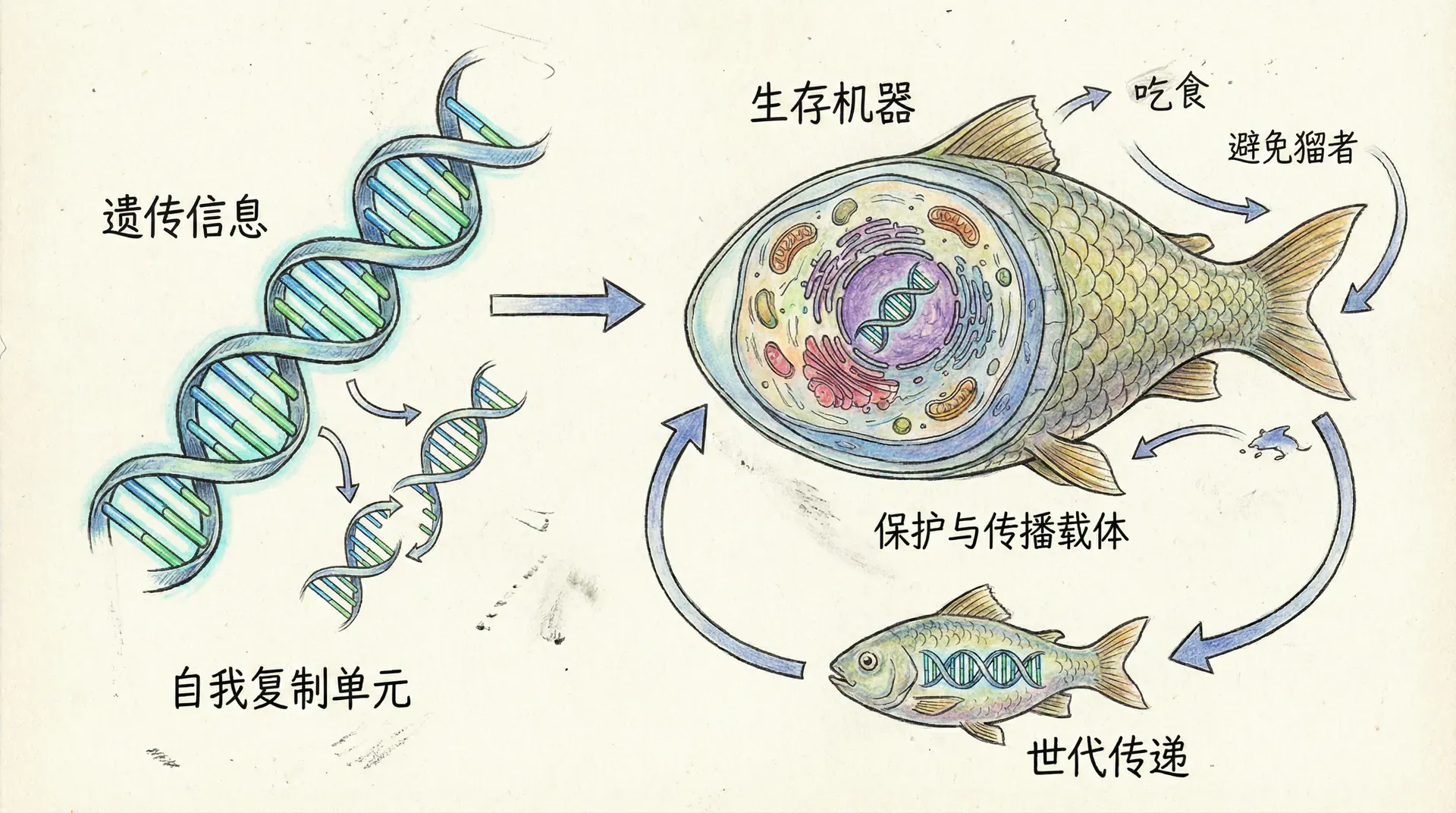
理解复制子概念
为了解决基因与个体之间的理论矛盾,我们需要引入“复制子”和“载体”的概念。复制子是指能够自我复制的基本单位,主要是DNA分子,但随着分子生物学的发展,我们还发现某些RNA分子、病毒甚至拟寄生因子都可以作为复制子。广义来看,任何能够信息自我拷贝并传播的遗传实体,都可以称为复制子。
而载体则是复制子构建的“生存机器”,通常是我们所说的生物个体。个体的层次可以多样,既可以是单细胞生物个体,也可以是如人类、银杏或鲸鱼一样庞杂的多细胞结构。更进一步,某些社会性动物种群(例如蚂蚁群体)从进化上也可以看作更高层级的载体。复制子和载体的区分,让我们能够用全新的角度来理解进化的核心动力和生物的复杂性。
可以用公司运营类比:DNA等复制子好比是“总公司的核心业务策略”,而生物个体则像分布在世界各地的“分公司”。每家分公司(个体)各自有不同的运营状况,但它们最终都服务于总公司的长远利益——即复制子最大范围的传播与存续。分公司不是目的,而是冲锋陷阵、为总部利益服务的执行机构。
这一思维框架帮助我们超越了以“物种”、“个体”为中心的传统生物学视角。我们能更好地解释一些看似“利他”或“自我牺牲”的行为本质上是复制子层面在“选择”最优传播路径。
载体的协调机制
那么,个体生物为什么能表现出如此统一和协同的行为?这里的奥秘在于,所有基因共享同样的“出口”——也就是精子或卵子等生殖细胞。无论是人还是银杏,从根本层面来说,体内的基因都只能通过受精卵这样的通道传给下一代。这仿佛所有运动员都必须通过同一个“终点”才能算作胜利,因而会更加自觉地团结合作,避免“内斗”带来的传递失败。
以中国的银杏树为例,这种植物可能存活几千年,一棵树体内有数万亿个细胞。尽管这些细胞可能会在不同生长期表现出功能差异(如叶、根、胚乳等),但其中的基因信息在生殖上是高度协调统一的,因为它们的最终命运是以同一颗种子、同一条“生命链”的形式进入未来的世代。
这种协调还体现在多细胞生物抗拒“内耗”的机制。例如,正常情况下,体内若有细胞“叛变”——也就是癌变——就是对“基因共同利益”的严重威胁。天然的细胞凋亡、免疫监控机制等,正是载体为稳定复制子传递而共同进化出来的内控体系。
个体存在的深层原因
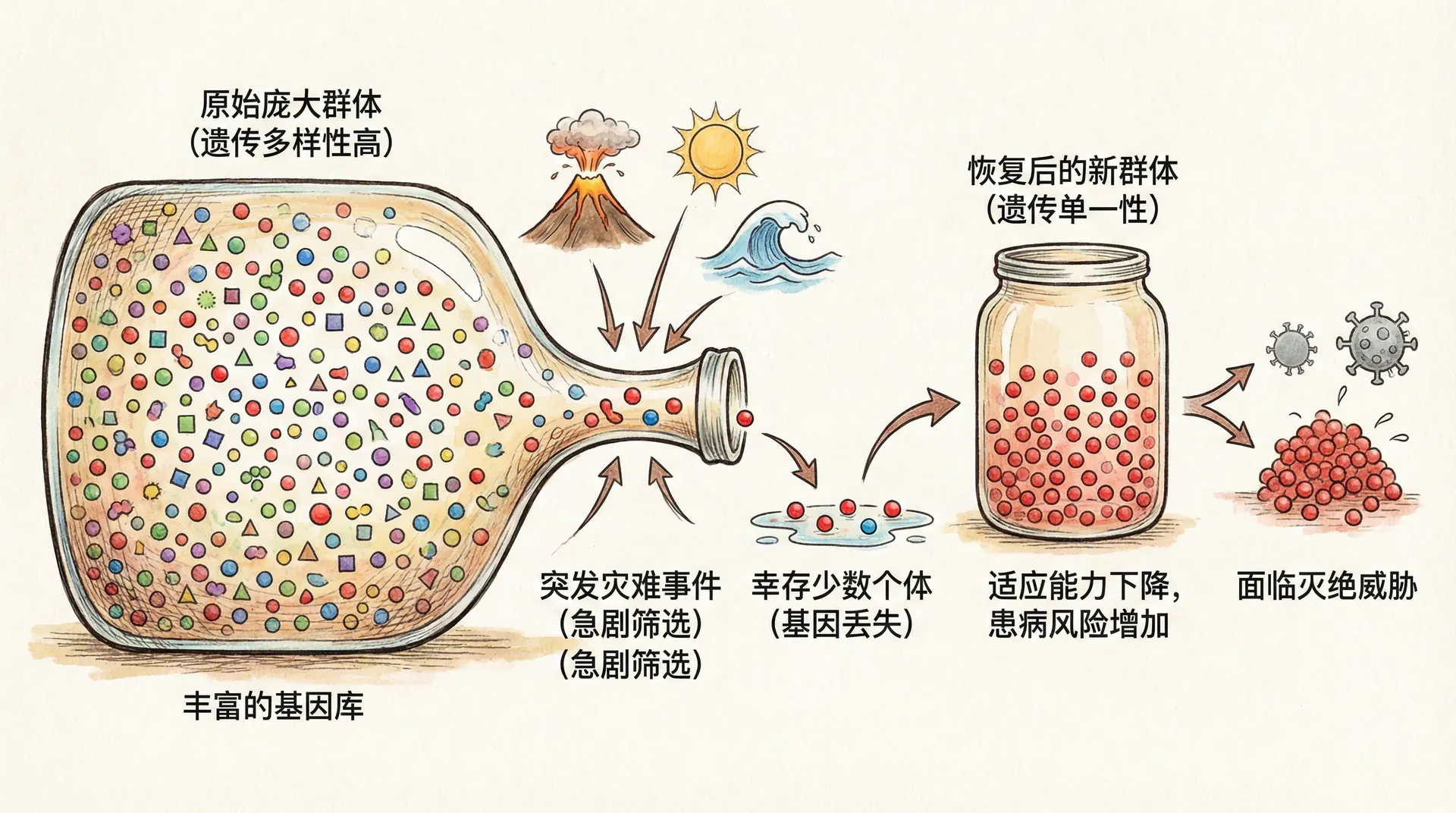
瓶颈效应的重要性
生物为何会进化出高度复杂的多细胞个体,而不是简单停留在单细胞阶段?关键因素是生命周期中的“瓶颈效应”——在每一代的更替周期,无论个体多么庞大、复杂,都必须回归到一个单一细胞(如受精卵)的起点。这个极小的细胞阶段使生命的信息得以精细整合、重组并筛选,也为进化中的优化提供了天然窗口。
如果没有这一“单细胞瓶颈”,许多遗传错误和体内累积的损伤会一代代继续下去,不利于适应复杂环境。例如,同样是多细胞生物,部分简单的群体性微生物(如部分菌落)不经历严格的单细胞重启,中间产生的无序扩增常常导致群体活力减退。相比之下,生命周期具有鲜明瓶颈的复杂生物(如哺乳动物、被子植物),每一代都像企业“年终大盘点”——剔除问题部门,保留最佳基因,为新一代奠定高质量起点。如下表所示,不同生物类群的生命周期瓶颈程度与复杂性之间有显著关系:
这种机制本质上为进化提供“清零机制”,让新一代不会被前代的全部缺陷和变异拖累。它也使得基因之间在下一代中有了“重新洗牌”的机会,增加了群体的适应和进步潜力。
发育程序的精确控制
正是依赖于生命周期中的瓶颈阶段,基因能够以一种“导演”的方式,对整个新生个体实施全面且有序的发育蓝图。从单细胞到复杂个体发育,像施工总指挥一样分阶段调配资源、定时表达指令,构筑独特而有序的生理构架。
以蝴蝶的变态发育为例,从卵到幼虫、蛹、成虫,不同分子信号和激素在精确时间点启动和关闭,少一步都可能导致尸体畸形或死亡。再比如,玉米籽粒的发育及成熟顺序,也有严格的遗传程序调控,确保每一粒种子不仅遗传可靠,还拥有足够的生理活力,见下表:
当代生物技术也在借鉴和利用这些发育程序。例如,诱导多能干细胞(iPSC)、再生医学和器官工程,正是利用了瓶颈-重启-重塑的逻辑,将人类细胞“按需归零”为全新用途。这不仅验证了发育瓶颈的重要性,也为疾病治疗和结构重建开启了新路径。
瓶颈效应构筑了生命世代更替的保障机制,确保基因和发育程序既有严格筛选又拥有创新空间。这一科学原理在农业育种、遗传疾病防治、生态保护等领域至关重要。
延伸表现型的实际意义
生态系统工程师
当基因的影响突破个体界限,便走向了“延伸表现型”。许多生物通过构建和改善环境,为自己及后代创造生存优势。比如中国水獭(与欧美河狸类似),在河流沿岸构筑巢穴和通道,不仅改善了自身的生存环境,还显著提升了周边湿地的生物多样性。这些“生态工程师”物种的基因通过行为影响环境,最终回馈自身。
生态系统工程师常见的代表如下:
这些生态系统工程行为不仅提升了环境多样性和生态稳定性,还常常成为其他物种赖以生存的关键基础。比如,珊瑚虫分泌碳酸钙造就的珊瑚礁,是无数海洋生物的庇护所。又如白蚁在非洲修建的大型蚁丘,土壤养分和通气极大提升,周边植被也更繁茂。
值得注意的是,有些物种的“工程”还有连锁正反馈,例如鸟类筑巢带动植物种子分布、水獭营巢提升湿地水质,整体优化了微生态结构。
农业中的应用价值
延伸表现型理论为农业生态设计提供了全新思路。传统上,育种和田间管理常只局限于作物自身性能,而忽视了基因对环境儿童的“溢出”影响。新一代农业模式则更加注重通过基因—环境联动,实现作物与生态系统的协同优化。
例如,棉花和玉米的不同品种根分泌物会吸引特异的土壤菌群,有的促进矿质养分释放,有的强化抗逆能力。实际应用中,科学家发现“功能型根分泌”品系能大幅减少化肥和农药使用,提升田间稳定性和可持续性。同时,不同耕作制度(如轮作、间作、覆盖作物)更能使不同作物品种间表现型互补,相互促进防虫和营养循环。例如:
- 蚕豆与小麦轮作:蚕豆固氮改善土壤,小麦减少病虫害;
- 水稻与鱼蟹共生:稻根分泌有益物质,吸引有益菌群,稻田生态更加稳健;
- 生物覆盖作物:如三叶草覆盖葡萄园,富集益虫和提高地表水分。
以下展示了延伸表现型理论推动的关键农业变革:
生态农业、智慧农业正是以延伸表现型为理论基础,不断试点和推广生态互作型田间模式,使作物成为整个生态网络中的枢纽而非“孤岛”。
进化的长远视角
基因竞争的复杂性
当我们将视野拓展到更大尺度,延伸表现型揭示了进化过程中的复杂多层次竞争与协作。基因不只是在个体内部“你死我活”,还通过“外包工程”影响环境、塑造生态网络,间接促进自身传播。例如:
- 某些放线菌分泌抗生素,占据土壤空间并遏制竞争者,有益菌群则更易生存下来(微生物药物博弈)。
- 蜜蜂授粉行为促成植物授粉繁衍,蜜蜂与植物基因形成互惠联盟。
- 白蚁建丘,促使土壤动物群落进化出各类“工程师依赖型”特征。
例如,亚洲象频繁迁移会在林间开辟通道,促进种子散播,为包括自身在内的多种动物创造新资源点。象群的“延伸行为”结果不仅影响植被和动物区系,还反馈提高种群存活机会。
这种跨物种、跨生态层级的表现型远不止传统意义的“个体”,其实际效果会反过来深远改变基因与环境的互动格局。
未来研究方向
随着分子技术和生态监控工具的发展,我们对延伸表现型的理解正进入深水区。例如,通过CRISPR和生物追踪技术,科学家可精准操控植物气孔延展基因,观察作物水分利用与气候适应性的生态反馈;或者编辑土壤微生物代谢路径,分析其对作物生长与地力恢复的推动作用。未来,更多基因—生态—社会多级联动的现象将被发现和利用。
此外,跨学科学习已成趋势。生物信息分析、生态网络建模、远程环境监控等方法结合,为人类全面揭示基因如何远距离、跨尺度影响环境和社会行为打开了大门。未来的研究不仅要回答“基因做了什么”,更要解析“基因如何激起复杂连锁反应、并最终回馈于生物本身及其后代”。
新一代生物学研究正在着眼于基因与环境之间的深度互动,以及延伸表现型在多尺度生态中的作用。理解这种影响网络,将为农业、生态修复、公共健康等领域注入创新活力和科学决策依据。
重新认识生命的本质
延伸表现型理论让我们重新认识了生命的复杂性以及基因在生物世界中的深远影响力。基因不仅仅是个体内部的指挥官,更是生态系统的重要塑造者,它们的作用能够跨越个体边界,延伸并影响到更广泛的自然环境。在这一视角下,生物个体不再是孤立存在的单位,而是整个生态网络和基因关联体系中的一个重要节点。这种相互关系打破了物种和空间的界限,使整个生物圈成为一个紧密联系、有机共存的整体。
这种理论不仅具有重要的学术价值,更在实际领域中带来了深远的启示。无论是生物多样性保护、现代农业发展,还是医学研究与疾病防治,延伸表现型的思想都为我们提供了全新的分析框架和实践方法。理解基因在生态层面、群体层面乃至整个环境中的长距离作用,有助于我们更好地看待生物世界的相互关系及其精妙复杂。
只有认识到基因影响的广泛性和多层次,我们才能真正实现对生物多样性的保护和可持续发展的目标,这也是人与自然和谐共处的智慧基础。