微生物的生存智慧
在自然界中,微生物面临着各种各样的生存挑战。假如食物耗尽、温度骤变或者环境突然恶化时,这些肉眼看不见的小生命该如何应对?它们既不能像动物那样迁徙,也不能像植物那样通过深根获取更多养分。然而,经过亿万年的进化,微生物发展出了一套令人惊叹的生存策略。
例如,我国土壤中常见的枯草芽孢杆菌,在长江流域的稻田土壤、黄土高原的农田以及东北黑土地中都广泛存在。当养分充足时,它们快速繁殖;但当遭遇营养匮乏时,它们不会坐以待毙,而是启动一系列精妙的应对机制。有些个体会形成孢子——一种类似“休眠舱”的结构,可以在恶劣环境中存活数年甚至数十年。更令人意外的是,有些细菌个体会通过释放特殊物质杀死自己的“兄弟姐妹”,并利用它们释放出的养分延续生命,这种现象被称为“同类相食”。
微生物的生存策略不是单一的,而是根据环境压力的强度和持续时间,选择最适合的应对方式。这种灵活性正是它们能够在地球上存在数十亿年的关键。
这些生存策略背后隐藏着复杂的分子调控网络和种群动态规律。理解这些机制不仅能帮助我们认识微生物的生命本质,也对农业生产、环境保护和生物技术应用具有重要意义。比如,在我国北方的设施农业中,利用枯草芽孢杆菌制成的生物菌肥已经广泛应用于蔬菜大棚,帮助提高作物抗病能力。而要更好地利用这些微生物,我们首先需要理解它们的生存策略和相互作用方式。
微生物之间的关系网络
在自然环境中,微生物很少单独存在。就像人类社会中存在各种人际关系一样,微生物之间也形成了复杂的相互作用网络。这些关系直接影响着每个种群的生存和繁衍。
生态互动的基本类型
我们可以根据互动结果对双方的影响,将微生物间的关系分为几种主要类型。下表总结了这些互动类型及其特点:
在这些关系中,竞争是最为常见的一种。当环境资源有限时,不同微生物种群之间就会展开激烈的竞争。这种竞争可以发生在同一物种的不同个体之间(种内竞争),也可以发生在不同物种之间(种间竞争)。
种内竞争的极端形式
种内竞争有时会演变为极端的形式——同类相食。这听起来似乎违背了"生物会保护同类"的直觉,但从进化的角度看,这恰恰是一种高效的生存策略。
以长江流域水稻种植区的厌氧环境为例。在水淹土壤深层,硫酸盐还原菌和产甲烷菌都需要利用有机物作为碳源和能源。研究发现,在靠近土壤表层的区域,硫酸盐还原菌占据优势;而在更深的厌氧层,产甲烷菌则占据主导地位。这种分层现象反映了两类微生物之间对相同底物的激烈竞争,竞争力较强的硫酸盐还原菌优先利用了上层的资源。
在实验室培养条件下观察发现,当营养物质逐渐耗尽时,枯草芽孢杆菌种群中约有30-40%的个体会死亡,而剩余的60-70%个体则通过利用死亡细菌释放的营养物质维持生存并延迟孢子形成。这种策略使种群整体在营养匮乏时期的存活率提高了数倍。
同类相食的分子机制
枯草芽孢杆菌的生存抉择
枯草芽孢杆菌是我国农田土壤中的常见微生物,在华北平原的小麦田、长江中下游的稻田以及珠江三角洲的蔬菜地中都能找到它们的身影。这种细菌具有一个非凡的能力:根据环境条件在不同生理状态间切换。
当细菌从生长旺盛期进入稳定期时,它们会按照时间顺序经历几个不同的发育阶段。首先是感受态发育,细胞能够从环境中吸收DNA片段并整合到自己的基因组中,这相当于“学习”其他细菌的遗传信息。接着是同类相食阶段,部分细菌会杀死其他细菌来获取营养。如果环境持续恶化,最终细菌会进入孢子形成阶段,将自己包裹在坚固的“保护壳”中度过难关。
致死因子的产生与调控
同类相食过程的核心是一个名为Spo0A的调控蛋白。这个蛋白就像一个“决策者”,根据环境信号决定细胞的命运。在营养缺乏的压力下,种群中只有一部分细胞会激活Spo0A蛋白,而另一部分细胞则保持Spo0A非活性状态。
在Spo0A活性较高的细胞中,会启动skf操纵子的表达。这个基因簇包含多个基因:skfA基因负责编码一种杀伤性肽类物质(致死因子),而skfE和skfF基因则共同组成一个“泵出系统”,能够将致死因子主动排出细胞外,同时保护自己不受致死因子的伤害。可以把这个系统比作“既能制造武器,又配备了防护盔甲”。
与此同时,另一个重要的基因簇sdp操纵子也被激活。sdpC基因产生的信号蛋白能够调控代谢相关基因的表达。它会促进yvbA转录因子的产生,后者进一步激活ATP合成酶基因(atp操纵子)和脂质分解酶基因(yusLKG操纵子)。这套系统的作用是帮助细胞从死亡同类释放的营养物质中快速提取能量,从而延迟进入“休眠”的孢子状态。
不同营养条件下细菌种群数量的变化趋势:
从图中可以看出,在营养缺乏条件下,Spo0A非活性细胞群体数量急剧下降,而Spo0A活性细胞则能够维持相对稳定的数量。这种“牺牲一部分、保存一部分”的策略,从种群整体的角度看是高效的。
保护与攻击的双重系统
有趣的是,即使是Spo0A非活性的细胞也不是完全没有防御能力。sdpC信号分子具有扩散性,它不仅作用于产生它的细胞,也能被邻近的Spo0A非活性细胞感知。在这些细胞中,sdpC会抑制σ因子相关基因的表达。σ因子是细菌转录过程中的重要蛋白,参与抗生素抗性和解毒反应。因此,缺乏σ因子保护的细胞更容易被致死因子杀死。
这种机制确保了种群中存在明确的“猎手”和“猎物”角色分化,避免了所有细胞都试图攻击对方而导致的混乱局面。从某种意义上说,这是一种通过分子信号实现的“社会分工”。
孢子形成
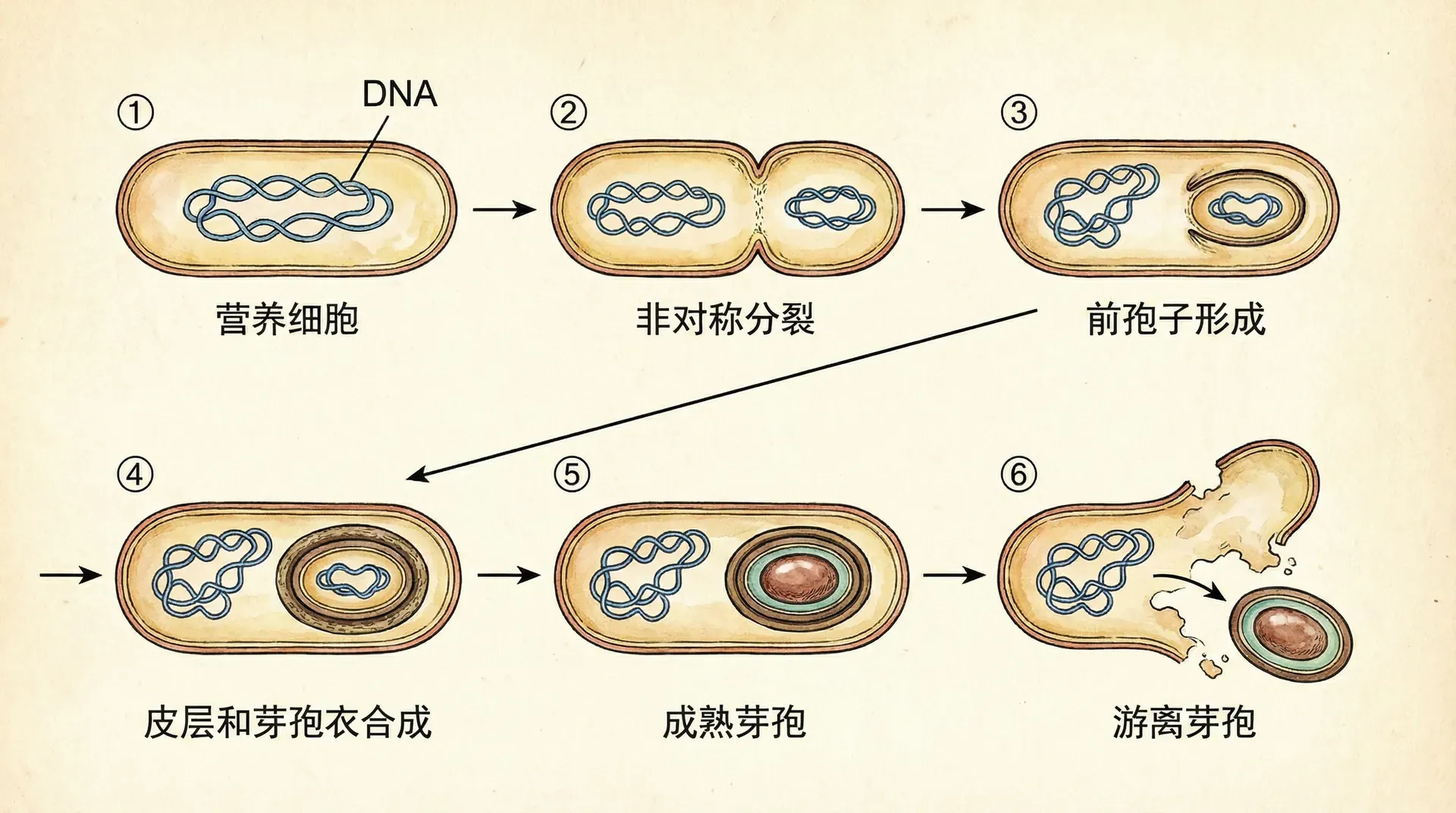
孢子的形成过程
当营养缺乏持续存在,同类相食也无法提供足够的能量时,枯草芽孢杆菌会启动孢子形成程序。这是一个高度复杂的发育过程,涉及超过100个基因的精确调控。
孢子形成可以分为几个连续的阶段。整个过程大约需要8小时完成。初始阶段,细胞会进行不对称分裂,形成两个大小不同的隔室:较大的母细胞和较小的前孢子。母细胞的作用是为前孢子的发育提供营养和保护,自己最终会裂解死亡。在这个阶段,两个隔室中分别激活不同的σ因子:母细胞中是σE,前孢子中是σF。
接下来,母细胞会像“吞噬”一样将前孢子完全包裹进来,形成双层膜结构。此时前孢子中的σG因子被激活,而母细胞中则激活σK因子。随后,母细胞开始合成皮层物质(一层特殊的肽聚糖结构)和外壳蛋白。皮层位于两层膜之间,而外壳则在前孢子表面组装,这些结构赋予孢子极强的抗性。
最后,母细胞裂解,成熟的孢子释放到环境中。这个孢子可以抵抗高温(能耐受80-90°C的温度)、紫外线辐射、干燥和许多化学物质。在我国新疆地区的干旱土壤中,或者在青藏高原的强紫外线环境下,都能发现处于孢子状态的枯草芽孢杆菌。当环境条件改善、重新出现适宜的温度和水分时,孢子会萌发,恢复为正常的营养细胞,继续生长繁殖。
孢子形成的调控网络
孢子形成的启动同样依赖于Spo0A蛋白。但与同类相食不同的是,孢子形成需要Spo0A达到更高的活性水平。这个蛋白通过磷酸化修饰被激活,磷酸基团通过一个复杂的“磷酸接力”系统传递。
这个磷酸接力系统包括五个组氨酸激酶(KinA、KinB、KinC、KinD和KinE)和两个中间传递蛋白(Spo0F和Spo0B)。不同的激酶响应不同的环境信号:有的感知营养物质浓度,有的感知细胞密度,还有的感知氧气水平。当多个激酶同时活跃时,磷酸基团像接力棒一样依次传递:激酶→Spo0F→Spo0B→Spo0A,最终使Spo0A被充分磷酸化,达到启动孢子形成所需的活性阈值。
下面的表格总结了孢子形成过程中的主要阶段和标志性特征:
检测孢子的实验方法
在实际研究和应用中,我们经常需要检测培养物中的孢子数量。常用的方法有两种。
-
染色法:孢子具有特殊的细胞壁结构,常规染色剂难以穿透。实验中使用孔雀石绿作为主要染料,通过加热促进染料渗入孢子内部。一旦染上颜色,孢子不容易脱色,会显示绿色。而营养细胞则用番红进行复染,呈现红色。在显微镜下观察,绿色的圆形或椭圆形结构就是孢子。这种方法在我国各地的微生物实验室中广泛使用,因为操作简单、成本低廉。
-
热处理培养法:利用孢子耐热的特性,将样品在90-100°C水浴中加热10分钟,这个温度足以杀死营养细胞,但孢子能够存活。然后将处理后的样品涂布在培养基上培养24小时,长出的菌落数量就代表原样品中的孢子数量。这种方法更加准确,常用于工业生产中的质量控制,比如在生产枯草芽孢杆菌生物农药时监控孢子含量。
种群动态的数学描述
竞争与互作的数学模型
要深入理解微生物种群的变化规律,仅仅进行实验观察是不够的,我们还需要用数学语言来描述和预测这些动态过程。
最经典的种群互作模型是Lotka-Volterra方程。这个模型最初用于描述捕食者-猎物关系,但也适用于分析同类相食等现象。对于两个相互作用的种群,其动态可以用一对微分方程来描述:
在这组方程中,代表被捕食者(或被攻击者)的数量密度,代表捕食者(或攻击者)的数量密度。参数是被捕食者在无捕食压力下的内在增长率,是捕食作用系数(反映捕食者找到并杀死猎物的效率),是捕食者的转化效率(表示捕食者从每个猎物中获得的繁殖收益),是捕食者的自然死亡率。
将这个框架应用于枯草芽孢杆菌的同类相食现象时,我们需要做一些调整。因为“捕食者”和“猎物”实际上是同一物种的不同生理状态个体,它们之间的互作更加复杂。
枯草芽孢杆菌种群的动态振荡
在无营养培养基中培养枯草芽孢杆菌时,观察到一个有趣的现象:活细胞数量不是单调下降的,而是呈现出周期性的波动。这种振荡模式可以用延迟微分方程来描述。
振荡产生的原因在于同类相食过程的时间延迟效应。当Spo0A活性细胞开始产生致死因子时,需要一段时间才能积累到足够杀死周围细胞的浓度。细胞被杀死后,其内容物释放到环境中,又需要时间才能被存活细胞摄取利用。这个摄取利用的过程还需要时间来转化为新的细胞物质。每个环节都存在时间滞后。
研究发现,振荡的幅度和周期与温度密切相关。在20-35°C的适宜温度范围内,能观察到明显的振荡;而当温度超过40°C时,死亡速率过快且致死因子不稳定,振荡现象消失,种群呈现简单的指数衰减。在低于20°C时,所有生化反应速率都显著降低,振荡幅度变得很小。
下方展示了不同温度下种群数量的变化模式:
从这个图表可以看出,在25°C时细胞数量呈现明显的波动(同类相食引起的振荡),在37°C时振荡幅度减小但仍然存在,而在42°C高温条件下振荡消失,表现为持续下降的趋势。
营养物质对动态的影响
同类相食行为受环境中残留营养物质的显著影响。当在无营养培养基中添加少量葡萄糖(碳源)或者氨基酸混合物(氮源)时,种群动态会发生改变。
实验发现,碳源的影响远大于氮源。添加葡萄糖后,同类相食的启动时间明显延迟,延迟的时长与葡萄糖浓度正相关。当葡萄糖浓度达到0.5%以上时,在观察期内(72小时)几乎不发生同类相食。这是因为细菌优先利用环境中的葡萄糖作为能源,只有当葡萄糖被消耗殆尽后,才会转而激活同类相食机制。
相比之下,添加等摩尔浓度的氮源(如酪蛋白胨)对同类相食启动时间的影响较小,通常只延迟3-6小时。这个差异揭示了一个重要的生理学原理:同类相食主要是为了获取能量(ATP),而不是为了获取氮元素用于合成蛋白质。换句话说,在营养匮乏时,细菌的首要需求是维持基本的代谢活动,而不是合成新的细胞物质。
下面信息对比了不同营养添加条件下的同类相食特征:
这个表格清楚地显示了碳源供应对延缓同类相食和孢子形成的显著效果。这一发现在实际应用中很有价值。例如,在发酵生产枯草芽孢杆菌生物制剂时,通过精确控制碳源的添加时间和浓度,可以最大化活菌数量并推迟孢子形成,从而提高产品质量。
捕食关系
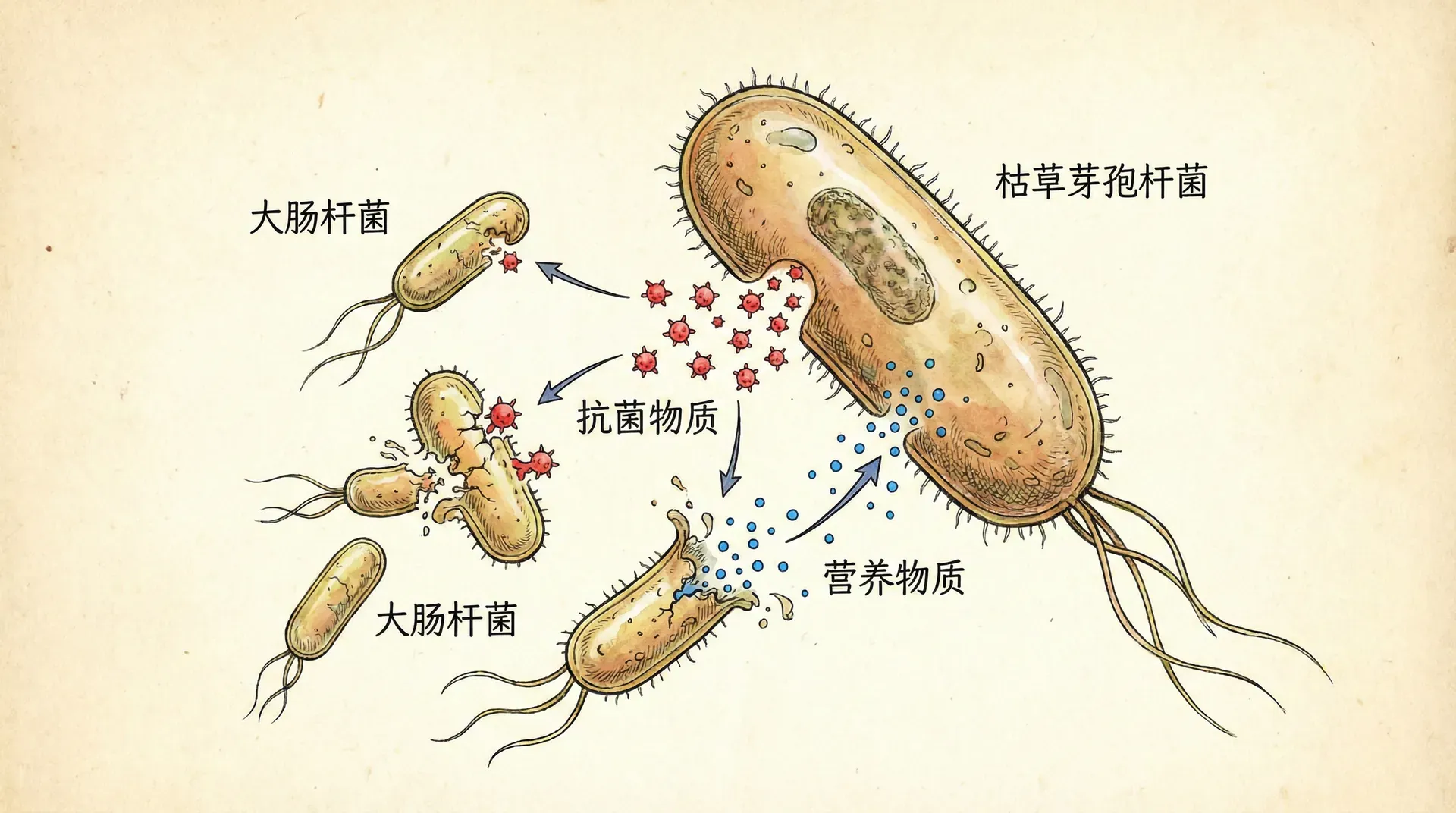
枯草芽孢杆菌对大肠杆菌的捕食
同类相食只是枯草芽孢杆菌应对营养匮乏的策略之一。当环境中存在其他种类的细菌时,枯草芽孢杆菌还会表现出跨物种的攻击行为——捕食作用。
在混合培养实验中,将枯草芽孢杆菌和大肠杆菌按一定比例混合后转移到无营养培养基中。大肠杆菌是我国水体和土壤环境中广泛分布的一种革兰氏阴性菌,也常作为粪便污染的指示生物。实验结果显示,在纯培养条件下,大肠杆菌在无营养培养基中的存活时间约为24-36小时。但在与枯草芽孢杆菌混合培养时,大肠杆菌的存活时间显著缩短,通常在12-18小时内数量就下降到检测限以下。
更有趣的是,枯草芽孢杆菌的活菌数在初期不但没有下降,反而有所增加。这表明它确实从杀死大肠杆菌的过程中获取了营养。使用活细胞计数法(MBRT法)监测表明,在混合培养的前24小时内,枯草芽孢杆菌的代谢活性维持在较高水平,远高于其纯培养对照组。
这种捕食行为的分子机制与同类相食有相似之处,但也存在差异。枯草芽孢杆菌产生的抗菌肽不仅对自己的非活性同类有效,对大肠杆菌等革兰氏阴性菌也具有杀伤作用。实际上,枯草芽孢杆菌能产生多种抗菌物质,包括表面活性素、伊枯草菌素等,这些物质对不同类型的微生物具有选择性毒性。
混合种群的动态平衡
在自然环境中,多种微生物共存形成复杂的群落结构。以我国南方稻田土壤为例,在5-10厘米深度的土层中,同时存在着枯草芽孢杆菌、大肠杆菌、假单胞菌、放线菌等多种微生物。它们之间的相互作用共同决定了群落的组成和功能。
当土壤经历淹水-排水的循环时,氧气供应发生剧烈波动,部分厌氧或兼性厌氧菌会快速生长。此时枯草芽孢杆菌等好氧菌处于劣势。但当土壤重新暴露于空气中,枯草芽孢杆菌能够快速从孢子状态复苏,并通过产生抗菌物质抑制其他竞争者,重新建立种群优势。
这种动态平衡可以用扩展的Lotka-Volterra竞争模型来描述。对于包含n个物种的群落,每个物种i的种群密度随时间的变化可以用微分方程来表示。该方程包含物种的内在增长率、环境容纳量以及物种间的竞争系数。竞争系数反映了不同物种对资源的竞争能力:当系数大于1时,表示一个物种对资源的竞争能力强于另一个物种;当系数小于1时则相反。
在枯草芽孢杆菌和大肠杆菌的系统中,由于存在明显的捕食关系而不仅仅是资源竞争,需要在模型中加入捕食项,使其更接近捕食者-猎物模型的形式。参数的估计需要通过大量实验数据拟合获得。
以下是枯草芽孢杆菌和大肠杆菌在混合培养中的种群动态:
图表清晰显示,大肠杆菌数量在混合培养中快速下降,而枯草芽孢杆菌则经历了一个先增长后缓慢下降的过程。这种模式与典型的捕食者-猎物动态一致:捕食者在猎物充足时数量增加,当猎物耗尽后自身也开始衰减。
环境因子对生存策略的调控
温度的影响
温度是影响微生物生理活动的关键环境因子。我国幅员辽阔,从黑龙江的寒冷黑土地到海南的热带土壤,温度差异可达50°C以上。枯草芽孢杆菌的同类相食行为对温度变化高度敏感。
在15-35°C的范围内,温度升高会加快同类相食的启动速度。这是因为温度影响了多个关键过程的速率:致死因子的合成速率、其在环境中的扩散速率、对敏感细胞的杀伤速率,以及存活细胞利用释放营养物质的代谢速率。在数学模型中,这些速率常数都遵循Arrhenius方程,即:
其中是温度下的速率常数,是指前因子,是活化能,是气体常数。不同过程的活化能不同,导致它们对温度的响应敏感度也不同。
当温度超过40°C时,情况发生了质的变化。在这个温度下,致死因子的产生受到抑制(可能是因为相关酶的稳定性下降),同时细胞的热损伤速率急剧上升。结果是同类相食现象基本消失,种群表现为简单的热致死过程。这种现象在我国西北地区的夏季表层土壤(温度可达45°C以上)中可能具有实际意义。
在低温端(低于15°C),所有生化反应速率都显著降低。虽然同类相食机制仍然存在,但进程非常缓慢,细胞数量变化的幅度很小。在我国东北地区的春季和秋季,当土壤温度在10°C左右时,枯草芽孢杆菌主要以孢子形式存在,营养体的代谢活动处于最低水平。
初始细胞密度的影响
种群的初始密度是另一个重要因素。这涉及到微生物的“群体感应”机制——细菌能够感知周围同类细胞的密度,并据此调整自己的行为。
实验表明,当初始接种量低于 CFU/mL时,几乎观察不到同类相食现象。这是因为致死因子是分泌到环境中的,需要达到一定浓度才能有效杀死周围的细胞。在细胞密度很低时,即使Spo0A活性细胞产生致死因子,它在环境中迅速被稀释,无法积累到有效浓度。这些孤立的细胞会直接进入孢子形成程序,而不经历同类相食阶段。
当初始密度在到 CFU/mL范围内时,同类相食表现典型。密度越高,进入同类相食阶段的时间越早,振荡的幅度也越大。这是因为高密度意味着细胞之间的距离更近,化学信号的交流更有效,致死因子更容易达到致死浓度。
下表总结了不同初始密度下的种群行为特征:
这个结果对理解自然环境中的微生物行为很有启发。在营养丰富的环境(如富含有机质的堆肥或粪肥)中,细菌密度通常很高,同类相食可能是常见现象;而在贫瘠的土壤中,细菌分布稀疏,个体细胞更倾向于独立应对压力。
研究方法与技术
活细胞计数的挑战
准确测定混合培养体系中不同微生物的活细胞数量一直是微生物学研究的技术难点。传统的平板计数法虽然可靠,但操作繁琐、耗时长(通常需要24-48小时),而且难以区分处于不同生理状态的细胞。
近年来发展的亚甲基蓝还原试验(MBRT)提供了一种快速、有效的替代方法。这个方法的原理基于活细胞的呼吸代谢活性。亚甲基蓝是一种氧化还原指示剂,呈蓝色。当它被细菌的脱氢酶还原后,转变为无色的还原型亚甲基蓝。还原速度与细菌的代谢活性和数量成正比。
在实际操作中,将一定量的亚甲基蓝溶液加入待测样品,混匀后置于恒温培养箱中,记录溶液从蓝色完全褪色所需的时间。通过与标准曲线对比,可以推算出样品中的活细菌数量。这个方法特别适合用于监测发酵过程和快速评估抗菌剂的效果。在我国的乳制品工业中,MBRT已经被标准化为检测原料乳质量的常规方法。
分子生物学检测手段
要深入理解同类相食的分子机制,需要检测特定基因的表达情况。现代分子生物学技术为此提供了多种工具。
实时荧光定量PCR(qPCR)技术可以精确测定特定基因mRNA的数量,从而反映基因的转录水平。例如,通过检测skfA、sdpC、spo0A等关键基因的表达量变化,可以追踪同类相食和孢子形成程序的启动时间和强度。这项技术在我国各大学和研究所的分子生物学实验室中已经普及,成本也逐年下降。
蛋白质印迹(Western blot)技术则用于检测特定蛋白质的含量。通过特异性抗体识别目标蛋白(如Spo0A、σ因子等),可以直接观察调控网络在蛋白质水平的动态变化。结合磷酸化特异性抗体,还能区分Spo0A的磷酸化状态,这对理解其激活过程至关重要。
荧光报告系统是另一个强大的工具。通过基因工程手段,将绿色荧光蛋白(GFP)或其他荧光蛋白的基因融合到目标基因的启动子后面。当目标基因被转录时,荧光蛋白也同时表达,使细胞发出荧光。使用流式细胞仪或荧光显微镜,可以在单细胞水平实时追踪基因表达的异质性。这种方法揭示了种群中不同个体的Spo0A活性确实存在巨大差异,证实了“异质性分工”的假说。
环境样品的采集与分析
要将实验室发现应用于理解自然环境中的微生物行为,需要从实际环境中采集样品进行分析。以土壤样品为例,标准的采样流程包括以下几个步骤。
首先,根据研究目的选择代表性地点。如果研究农田生态系统,应该在田间选择多个采样点,避免边界效应和局部干扰。通常按照对角线或棋盘格方式布点,每个地块采集5-10个样本后混合。
采样深度根据研究对象而定。枯草芽孢杆菌主要分布在0-20厘米的土层中,因此通常采集表层(0-5厘米)、浅层(5-10厘米)和深层(10-20厘米)三个层次的样品。采样工具需要经过灭菌处理,避免交叉污染。
样品采集后应尽快处理或在4°C条件下保存,保存时间不宜超过24小时。处理时先除去石块和植物残体,充分混匀后进行系列稀释,然后分别进行平板计数、生理生化测定和分子检测。
在我国一些地区开展的农田微生物多样性调查中,研究人员发现枯草芽孢杆菌在有机质含量较高的土壤(如黑土、水稻土)中的丰度显著高于贫瘠土壤(如红壤、荒漠土)。这与其生存策略相吻合:在有机质丰富的环境中,即使进入营养匮乏期,同类相食和捕食策略也更容易获得足够的营养。
应用前景与实践意义
生物防治技术
枯草芽孢杆菌的捕食和抗菌特性使其成为优秀的生物防治菌株。在我国,多个枯草芽孢杆菌菌株已经被开发成商业化的生物农药和生物肥料产品。
以防治黄瓜白粉病为例,这是我国北方设施蔬菜种植中的常见病害,传统上需要使用化学杀菌剂防治。现在,将特定的枯草芽孢杆菌菌株制成菌剂喷施在叶面上,能够有效抑制白粉病菌的生长。其机制包括多个方面:菌株在叶面定殖后竞争营养和空间,产生的抗菌肽直接抑制病原菌,还能诱导植物自身的防御反应。
在山东寿光的大棚蔬菜种植区,已有多家合作社采用枯草芽孢杆菌生物菌肥。农户反映,连续使用2-3季后,土壤传染病害的发生率明显下降,化肥使用量减少约20-30%,蔬菜产量和品质都有提升。这种生态友好的防治方式符合绿色农业的发展方向。
发酵工业优化
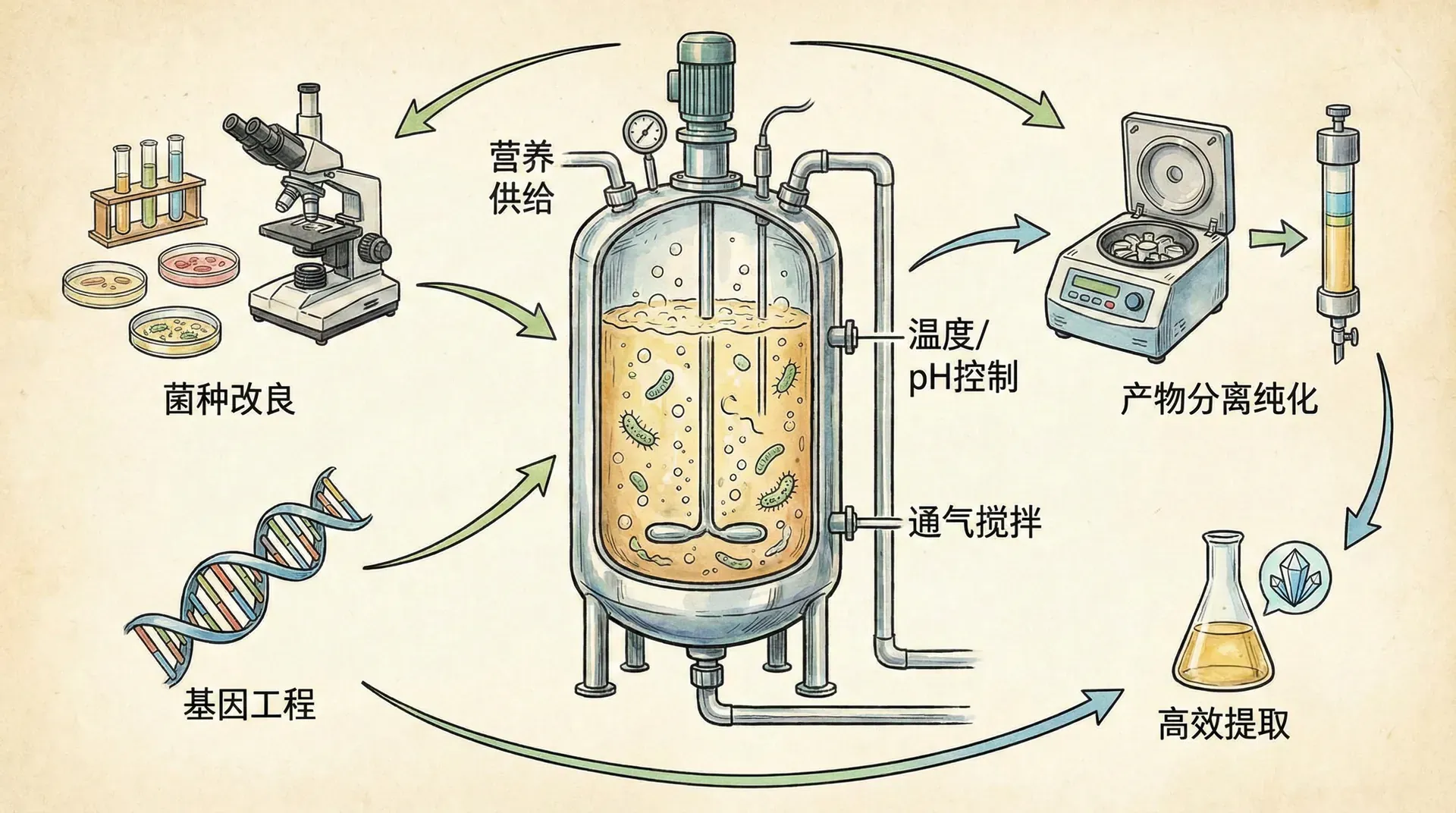
理解枯草芽孢杆菌的生存策略对优化工业发酵过程也有重要意义。在生产酶制剂、抗生素或其他代谢产物时,需要维持细菌在活跃的营养生长状态,避免其过早进入孢子形成程序。
根据前面讨论的营养效应,通过精确控制碳源的供应,可以有效延迟同类相食和孢子形成。一些先进的发酵工艺采用流加培养策略:不是一次性加入所有葡萄糖,而是根据在线监测的溶氧和pH值,少量多次地补充碳源,使其始终维持在一个较低但非零的水平。这种“饥饿边缘”的培养条件能够保持细菌的代谢活性,同时最大化目标产物的产量。
在我国的酶制剂生产企业(如集中在山东、江苏、河北等地的发酵工业园区),这些优化策略已经带来了显著的经济效益。某枯草芽孢杆菌α-淀粉酶生产工艺经过优化后,发酵周期从72小时缩短到48小时,酶活力提高了40%,每吨产品的成本降低约15%。
环境修复潜力
枯草芽孢杆菌对多种有机污染物具有降解能力,在环境生物修复中展现出应用潜力。它能产生多种胞外酶,包括蛋白酶、脂肪酶、纤维素酶等,这些酶可以分解复杂的有机化合物。
在我国南方的一些水体富营养化治理项目中,研究人员尝试投加枯草芽孢杆菌菌剂来加速有机污染物的降解。在浙江太湖流域的一个试验点,投加菌剂后水体中的化学需氧量(COD)在两周内下降了约35%,总氮和总磷也有明显降低。机制分析表明,枯草芽孢杆菌不仅直接降解有机物,还通过产生抗菌物质抑制了蓝藻的生长,从而减缓了富营养化进程。
在石油污染土壤的修复中,某些枯草芽孢杆菌菌株表现出降解多环芳烃的能力。在陕西延安地区的油田区,研究人员从石油污染土壤中筛选出高效降解菌株,经过实验室和中试规模的验证,这些菌株能够在3个月内将土壤中的总石油烃含量降低50%以上。目前相关技术正在申请专利并准备推广应用。
从微观机制到宏观现象
对枯草芽孢杆菌生存策略的研究揭示了微生物在不同层级的灵活适应性:从基因调控到细胞状态转变,再到种群动态乃至群落互作。其同类相食行为和孢子形成,实质上是应对环境压力的两种关键策略,前者提升了营养匮乏时的“群体”存活概率,后者则为长期恶劣环境提供了休眠保障。细菌能够根据外界条件灵活切换策略,其背后有复杂的遗传调控和生态动力学基础。数学模型和实验证据不断丰富我们对这些机制的认识,并指导发酵、环境修复等实际应用,强调环境变量对菌株生理和表现的影响。
目前,枯草芽孢杆菌已广泛应用于农业、工业与环境领域,未来在绿色农业、生态修复等方面还有更大潜力。利用微生物多样的生存策略,不仅拓展了基础研究的深度,也为可持续发展提供了创新思路和工具。
微生物的生存智慧告诉我们:多样性的策略才能适应复杂环境。系统思维与工程优化,将推动微生物学研究和应用继续前行。