理解生物复杂性
当你观察黄河滩涂上成群的候鸟迁徙、秦岭密林中金丝猴的灵巧动作,或是沙漠边缘胡杨林在风中屹立、热带雨林里蚂蚁王国的精密分工时,都会不由自主地产生敬畏:如此多元且高度有序的生命世界究竟是如何构建起来的?
无论是鲤鱼在江河中逆流而上,还是深海热泉旁顽强的微生物群落,或者极地雪地里企鹅育雏,这些生物纷繁的形态和极强的适应力,都不断刷新我们对“复杂性”的想象。与非生物——如岩石、河流、沙丘、月亮——相比,生物体不仅仅在组成和结构上复杂,更表现出信息处理、协调调节以及对环境变化的主动响应。
理解生命系统的复杂性与其起源和运行机制,是生物学的根本问题。它不仅关乎学术探索,更决定着我们对健康、生态保护以及未来生物技术的核心认知。
什么是复杂性?
复杂性的基本特征
要理解生物的特殊之处,我们首先需要明确什么是“复杂性”。复杂性并不仅仅体现在组成部分多,更关键的是这些部分之间如何协同分工、相互作用。复杂系统的本质在于:其结果远超各独立部分的简单叠加,常常表现出新颖的整体功能(涌现)。
让我们用一个比较来说明这个概念:
我们会发现,生物系统和高等级人工系统之所以被认为复杂,核心在于:它们的组成元素不仅多,而且结构严密、有分工、有信息处理能力和反馈调节机制。同时,还能根据环境变化主动调整和自我修复。
统计不可能性
真正的复杂性具有一个关键特征:它在统计学上极不可能通过纯粹的偶然产生。即使我们给予充足的时间和空间,单靠随机组合也极难获得类似生物体那样协作严密、具备精确功能的结构。为此,我们可以用简单的概率分析和图表来直观理解。
从图中可见,随着组件数量的增加,通过纯粹随机组合产生有功能的结构,其概率呈指数级下降。仅仅是十几个组件,就已极度罕见;而众多生物结构动辄涉及数百乃至上千种分子成分!这便解释了为什么复杂的生命体系无法简单归因于偶然事件。真实的生物复杂性往往需要累积的选择和历史性的优化过程。
复杂性的本质在于:它是一种在事先规定的标准下,极不可能通过随机过程产生的有序状态。这样的有序不仅仅是静态的排列,更关乎动态功能的精确达成。
从功能角度理解复杂性
生物系统的复杂性还体现在它们拥有高度明确、精密协作的功能目标。生命体能够精准地将各种原材料转化为能量,进行信号传递、环境感知、自我修复等任务,这些功能通过复杂网络的协调运行得以实现。相比之下,人工和物理系统虽也具备某些功能,但其效率与自适应能力往往远逊于生物体。
以下的图表将不同系统的维护成本与其功能效率进行对比——这能帮助我们理解为什么“复杂”并不等同于“低效”,反而是高效与适应性的基础:
从图中可见,生物系统通常能够以较低的维护能耗持续高效运行,这种优化是数亿年进化带来的独特优势。相比之下,人工系统常常依赖外部能量和技术人员维护,物理系统则缺乏自我调节与高效功能。生命的复杂性正蕴藏于其“结构-功能”协同优化之中,也是生命与非生命之间最本质的差异。
生命系统的特殊性质
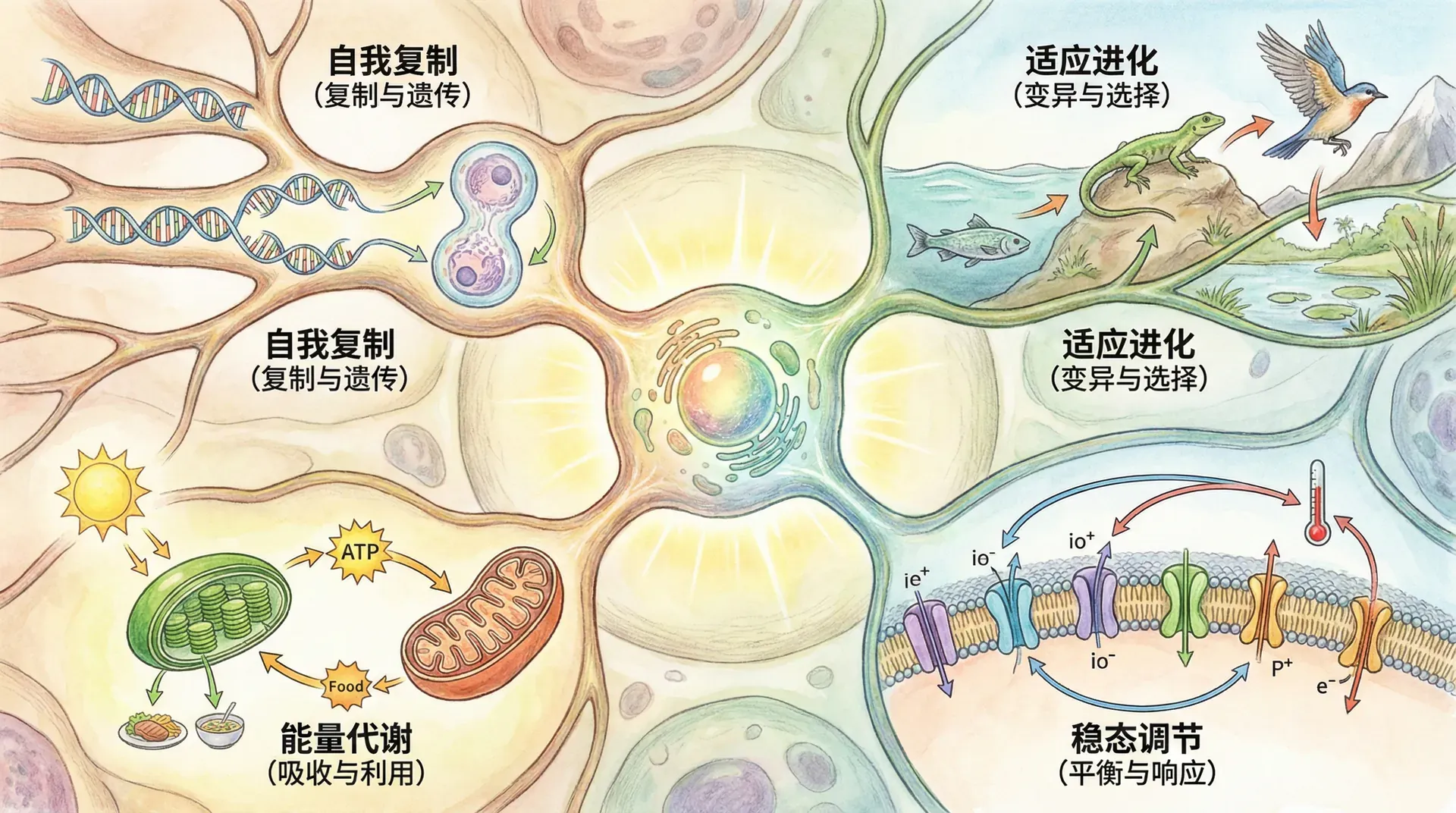
主动维持平衡
生物系统的一个核心特征,就是能够主动地维持和调节自身内部环境,使其即使在外部环境极端变化时,依然保持适合生存的稳定状态。这种能力被称为稳态(Homeostasis),是生命区别于非生命系统的关键之一。
在中国的生态系统中,这种主动维持平衡的现象表现得淋漓尽致:
-
胡杨树在塔克拉玛干沙漠边缘能够在极度干旱的条件下存活。胡杨树不仅拥有极为发达和深入地下的根系,以吸收深层稀少的水分;其叶子在炎热与干旱环境下还会变小甚至脱落,以减少水分蒸发。同时,它们体内含有较高的有机渗透物质,可以维持细胞不至于缺水萎缩。
-
藏羚羊能够在海拔4000米以上低氧、高寒的高原环境中正常生活。它们的血液中血红蛋白含量更高,能更有效地运输氧气;心肺系统具有特别的生理适应,甚至连细胞线粒体对氧利用能力都经过进化优化。研究还发现藏羚羊体内部分基因表达,在缺氧条件下会被上调,以增强生理反应。
-
竹子以惊人的速度生长,却能适应南北气候差异大、土壤贫瘠等环境条件。竹子的结构在生长过程中会自动调节,保证柔韧性与强度兼备;同时通过发达的地下茎网络,竹子群落能够在火灾、洪水后的较短时间内迅速恢复。
此外,许多中国本土生物也展现出主动调节体温、渗透压、酸碱度等多种稳态维持机制。例如,沙漠蜥蜴会在清晨吸收热量、正午躲入阴影调控体温;高原植物能够通过增加叶表蜡质层减少水分流失;长江中华鲟可以调节血液中的盐分,适应淡水与海水环境切换。
生物系统通过主动获取和利用外界能量,不断修复自身损伤、调节功能,维持高度有序。这与无生命系统趋向于热力学平衡与无序衰变正好相反,体现了生命“远离平衡态”的本质。
让我们用图表来展示不同系统在有序维持上的根本区别:
上图显示,“活细胞”等生物系统即便经历很长时间,依然能够维持高水平的有序度——这是因为其持续进行代谢、修复、调控等主动过程。而“死细胞”会因代谢停止而迅速走向无序,机器设备虽然通过维护可减缓磨损,但远不能像生命那样自我更新。
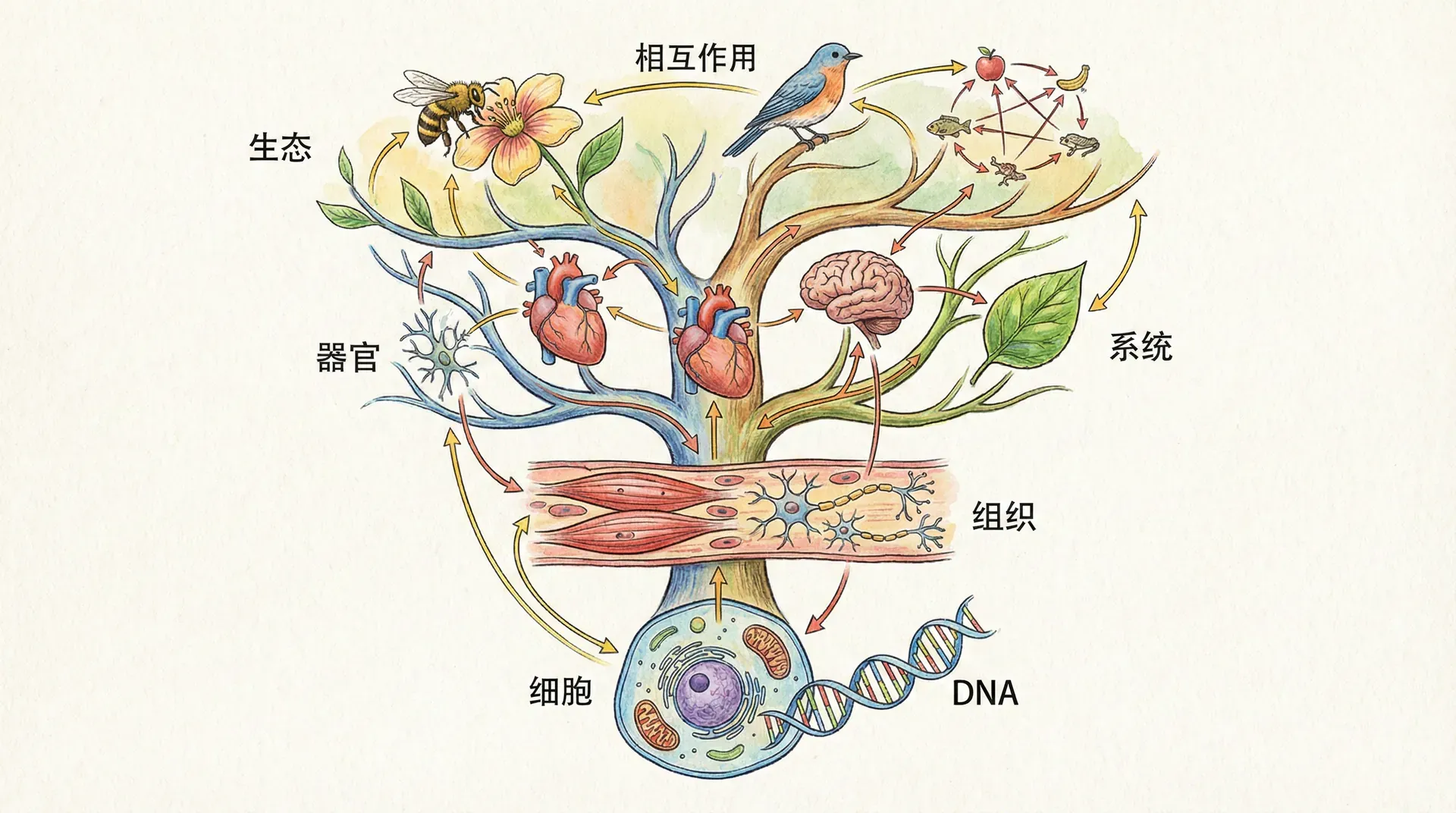
层次化的组织结构
生物系统的惊人复杂性,还体现在其多层级、嵌套式的组织结构。这种层次化不仅带来功能分工和效率提升,还增强了系统的适应力和演化潜能。
从分子到生态系统,每一层次都在中国自然界中有鲜活例证:
正是这种多层次、有机衔接的组织结构,让生物能同时兼顾局部与整体的功能,从微观分子操作到宏观生态调节,展现出令人赞叹的复杂之美与生命力。
设计的表象与真实机制
传统的设计论观点
当我们观察到生物世界的精巧结构乃至那些宛如“为功能而量身定制”的细致器官时,人们极易联想到背后存在着某种有智慧的造物主。这一思想,在哲学、宗教以及早期的自然史研究中,统称为“设计论证”。尤其是在达尔文之前的时代,大多数学者都会试图解释为何复杂的功能性仅靠自然过程难以实现。
假如有人在泰山之巅意外发现一块精美绝伦、雕纹细致的玉石,必然认为它必属工匠之手,而不是天然风化之果。这一判断有着鲜明的三重依据:
- 精确的几何形状 —— 自然风化形成规整几何的概率微乎其微
- 复杂的纹理 —— 丰富的装饰性图案极难靠自然作用随机生成
- 明显的功能性 —— 例如玉石作为馈赠、祭祀或权力象征之意
这样的推理模式沿用到生物界,人们自然会问:如此完美的结构(如鸟翼、鱼鳍、人眼和蜂巢),莫非不也是“人工计划”与有意设计的结果吗?
传统的设计论主张,诸如鸟类翅膀、鱼类鳍、甚至人眼这样高度复杂而具备特定功能的结构,必然需要一位全知全能的设计师才能形成,否则如此协调与高效难以由盲目过程导致。
历史上,这一观点广泛影响着科学与宗教的讨论。例如,古希腊哲学家柏拉图、亚里士多德、以及中世纪的基督教神学家和中国古代的“天工开物”思想,都不同程度地将生物的复杂性归因于“天意”或“自然之神”的安排。
达尔文革命
19世纪中叶,查尔斯·达尔文提出“进化论”及“自然选择理论”,彻底改变了我们对于生物复杂性的认知。达尔文革命的核心意义在于:生物的复杂性和适应性,完全可以由无意识、无目的的自然过程—尤其是变异与选择—逐渐积累演化出来,无需依赖“智能设计者”或超自然力量。
让我们用一个模拟图来帮助理解这个过程下的演化动力:
该图模拟了两种不同演化路径:一条是“有利突变累积”,即每一代有益变异经自然选择被保留下来,其适应性不断提升;另一条则是“纯随机变化”,没有选择压力时系统的变化基本无序、不积累。
进化论解释复杂结构的四个关键环节:
变异 —— 每一代都会因为基因突变、重组等机制积累大量微小差异。
选择 —— 环境“筛选”出适应力更强的个体,劣势个体被淘汰,有优势的基因得以扩散。
遗传 —— 有利的遗传变异通过繁殖传递给后代。
累积 —— 长期作用下,细微变化汇聚成重大的结构和功能跃迁。
需要强调的是,这一过程既不是随机的(不是“骰子丢出来的结构”),也没有预先设定的目标,而是通过长期积累中的“差之毫厘,失之千里”式筛选最终产出适应性极高的结构。
生物进化的案例
在中国丰富的生物多样性中,我们能找到许多用进化解释而非“特殊设计”更为自然的例子:
大熊猫的拇指
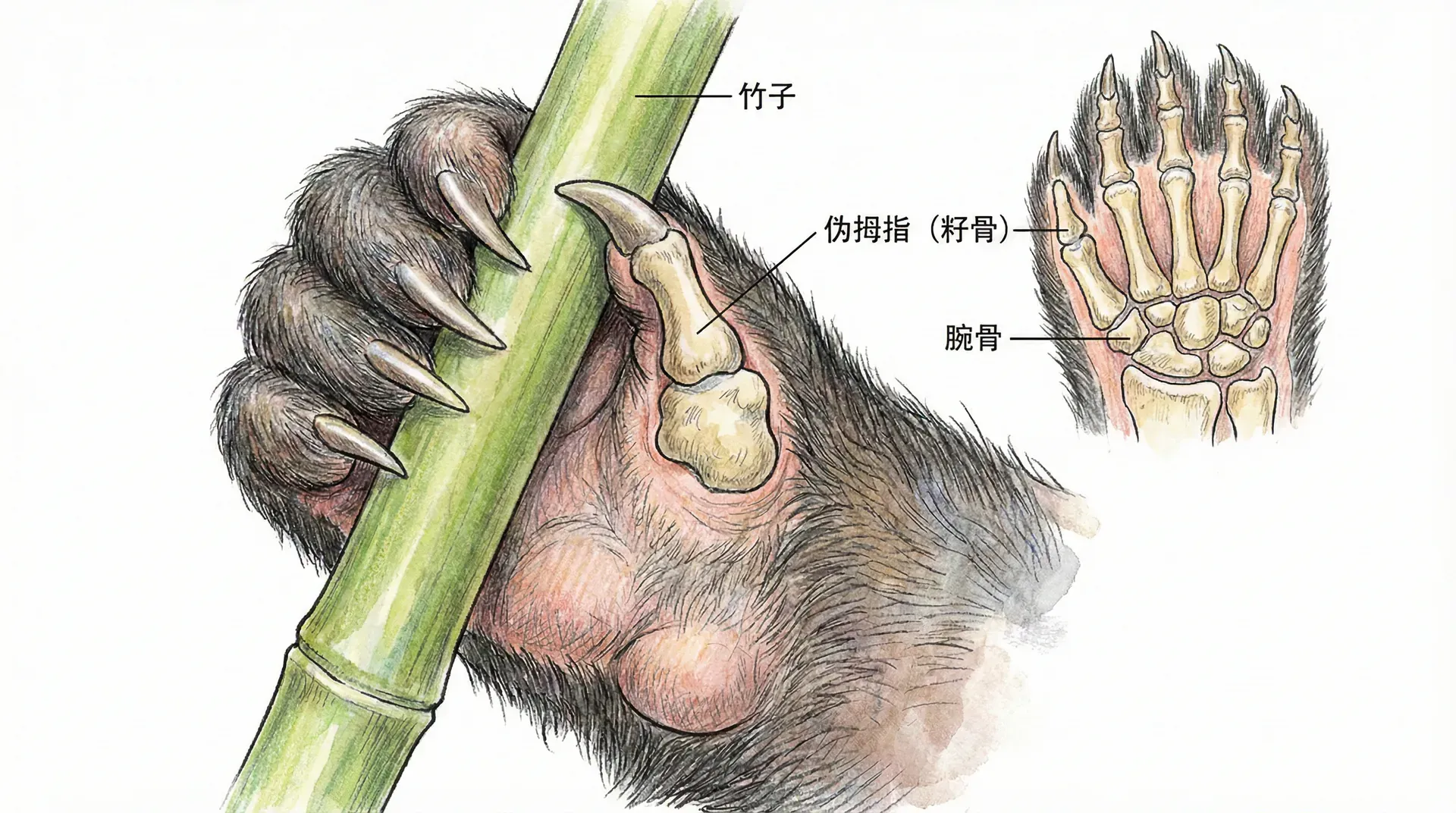
大熊猫的“伪拇指”可以说是达尔文主义下典型的“兼容性进化”案例。最初,祖先的腕骨仅仅用于行走和支撑;但随着饮食习性的转变,为了更好地抓握竹子,这一腕骨逐步演化成类似于拇指的结构。这并非“设计者”有意添加了一个拇指,而是骨骼逐步适应抓握动作的需求,最终形成帮助采食竹子的“伪拇指”。
- 进化亮点:结构并不完美,例如大熊猫的伪拇指比人类拇指灵活性差;但它已极大增进了采食竹子的效率。
- 科学意义:说明复杂器官可由“现成架构”微调演变,无需推断“专项创造”。
青藏高原生物的高原适应
青藏高原地处世界屋脊,无论是人类还是动物都面临着极低的氧气浓度。藏族人群与高原特有动物(如藏羚羊、雪豹等)分别通过遗传和生理机制适应极端环境:
- 藏族人群长期生活在高原,出现更高的红细胞密度、更强的呼吸及循环功能,部分原因在于血红蛋白结构的基因突变,有利于携氧能力增强;
- 与此同时,平原来的游客虽然短期能够通过“生理调节”提升红细胞数量,但远不及世世代代演化而来的“遗传适应”;
- 高原动物则往往拥有更加极端的生理适应,如血液携氧蛋白的特殊分子构造。
这些案例表明:生物复杂性不仅可以在达尔文进化论框架下清晰解释,而且不再需要假定有超自然的设计者。复杂结构和适应性源于亿万代的变异、筛选与积累,其“精巧”正是自然过程长期雕刻的结晶。中国生物多样性,也为进化理论提供了丰富的实证土壤。
理解复杂系统的方法
层次化还原论
要想真正理解生物的复杂性,我们需要引入“层次化”的视角。这就好比分析一座复杂的楼宇,既要看到整体结构,又要拆解到每一砖一瓦。层次化还原论是一种重要科学方法,强调我们可以通过分层理解和研究,把大问题分解为可管理、可研究的小部分。它要求我们既关注整体功能,又能深入到细节,从而实现“见树亦见林”。
以研究大熊猫的消化系统为例,这一系统本身就是多层次彼此嵌套、复杂交互的产物。科学家通常按从宏观到微观的顺序分层开展研究,每一层次都揭示了特定的信息和规律:
通过从整体到分子的多层次研究,我们不仅能理解大熊猫如何消化竹子,还可以揭示其进化如何优化了各个环节。例如,分子生物学发现大熊猫的消化酶基因有独特的变异,有助于适应高纤维饮食;而解剖学则说明其消化道容量与普通肉食动物截然不同。
层次化还原论并非只关注局部细节,而是帮助我们理解:复杂系统之所以能高效运行,正是在不同层次的协同作用下实现的。
不同层次并不是孤立存在:分子水平的微小变异可能会在细胞水平累积影响,从而改变组织功能、最终影响整体表现。反过来,外部环境的变化也可能通过整体层应激,从上而下地影响基因表达与分子机制。这种“上下贯通”的动态贯穿着整个生命科学研究,也是理解复杂性的关键。
从简单到复杂的解释路径
科学的目标之一,是能够将复杂现象还原为一系列可以理解、可预测的简单原理。真正深入的科学解释,就是从基础出发,逐步构建到更复杂的层级。
下方图表说明这种“自下而上”的解释模式:
每一个层次的原理都建立在更基础层次的原理之上,但同时具有该层次特有的规律和特征。比如生物学的“自然选择”,虽然根植于物理和化学定律,但却展现出全新的复杂性和创造力。
复杂系统的科学解释
以“雪豹如何抓捕藏羚羊”为例:
- 物理层面 解释奔跑与跳跃的力学过程;
- 化学层面 涉及能量的获取和消耗;
- 生物学层面 分析猎物追踪、捕猎行为的遗传机理和神经调控。
只有将所有层级的信息整合起来,才能获得全面的解释。这种多层级的、从简单到复杂的分析路径是自然科学研究的基石。
概率与必然性
生命的统计学奇迹
生命的出现和进化,乍看之下是在概率上极其不可能的事件。确实,如果我们把生命复杂结构的“最终形态”当作目标,计算其“一步达到”的概率,常常会得出震惊的、趋近于零的数字。不过,这种概率直觉其实误导了很多人。
让我们用抛硬币的例子来说明:如果我抛硬币10次,得到任何特定序列(比如“正正反正反反正反正正”)的概率都是1/1024。事后来看,每个序列都很“特殊”,仿佛是奇迹。但其实这只是我们事后的认知。如果我们事先指定一个特定序列,比如“连续10个正面”,那么得到它的确很罕见——但自然界的生命结构远不是单纯依赖一次达成,而是经历了无数代的选择和筛选。
同理,在生物界,我们看到的“复杂适应性结构”也许令人震惊,却常常是亿万年演化中累积筛选的结果,而非偶然间“预定目标”一次实现的事件。
累积选择的力量
进化的核心并不是要靠“幸运的一击”达成极端复杂度,而是依靠每一步小改良的积累和选择,逐步塑造适应性。这就是所谓“累积选择”的威力。
让我们以古代建筑为例,没有哪一座雄伟的宫殿是一次铸成的:
每一步都有较高实现概率,最终令人赞叹的宫殿,则是无数小步骤时间的积累之果。生物演化中的复杂器官、互作网络,也是如此。在每一次变异与筛选中,简单结构缓慢复杂化,因此最终“奇迹”并非概率的叠加难题,而是自然而然的历史产物。
理解进化的累积性,有助于我们摆脱“复杂即不可能”或“必有设计者”的误区。科学鼓励我们去寻找过程背后的合理机制,而不是止步于表象的不可思议。
自然界的“累积选择”例子
- 鸟类羽毛的演化:由鳞片逐步演化出简单绒毛、再发展成对飞行高度适应的复杂羽毛。
- 哺乳动物听骨:从爬行动物下颌骨逐渐转化为专门传声的中耳听小骨,过程可在化石中明确追踪。
- 植物光合作用体系:各种层次的蛋白复合体逐步叠加,实现了高效率能量转化。
总结
我们用科学的“解码”思维重新衡量了生物复杂性的本质。生物系统的惊人多样性与适应性,往往呈现出目的性、设计感。然而,这种“设计感”其实来源于无意识的自然过程,尤其是自然选择在极长时间尺度上的累积作用。
理解生物复杂性的关键在于:即使最复杂的结构,也可以通过简单规则的长期累积效应产生。这正是进化论揭示的自然之力,不需要超自然设计者也能解释世界的精妙。
这种科学理解极大拓展了我们的视野,也为生命科学、生态保护等领域奠定了坚实基础。在未来的学习中,我们将逐步分析具体生物,探索一切从分子到生态系统层面的适应机制,以及这些机制在现实世界中的巨大意义。
从秦岭的金丝猴到南海的珊瑚礁,从青藏高原的雪莲到东北的红松林,每一种生物都是伟大进化历程的活见证者。理解它们的复杂性,不只是学者的兴趣,也是现代社会保护多样性、可持续发展的必要前提。