生物复杂结构的渐进形成
当我们观察大熊猫那灵巧如“拇指”的特殊结构时,难免会好奇:如此精妙的“设计”究竟源自何处?事实上,这并非真正的手指,而是腕骨加厚变形、专为抓握竹子而优化的伪拇指,是竹食生存策略长期演化的结果。这类现象背后隐藏着共同的问题——高度复杂且契合需求的生物结构,究竟是怎样一步步从简单甚至“无关紧要”的前身逐渐演化而来的?它们似乎凝聚了巧夺天工的“设计智慧”,但实际上,自然界并不存在有意识的设计师,所有结构和功能的“奇迹”都诞生于漫长世代的自然选择。
类似的复杂结构不仅见于大熊猫,也广泛分布在中国许多代表性动物身上。例如,中华白海豚依赖精妙的回声定位系统在浑浊水域中探测猎物与障碍物;扬子鳄体表布满高灵敏感觉器官和复杂神经网络,即便极细微的水流变化也难以逃脱其感知;金丝猴则拥有应对高寒低氧的独特生理机制。所有这些高度复杂、契合环境的功能——其出现不仅是演化理论必须直面的难题,也是理解生命奇迹的核心。
理解渐进进化的关键问题
要解答这个谜团,仅靠直觉和“跳跃式”想象是远远不够的。我们需要系统地分析“复杂性起源”的全过程,从理论和实例层面去拆解渐进进化的运作原理。关于复杂结构的形成过程,科学家们提出了几个至关重要、互相关联的问题:
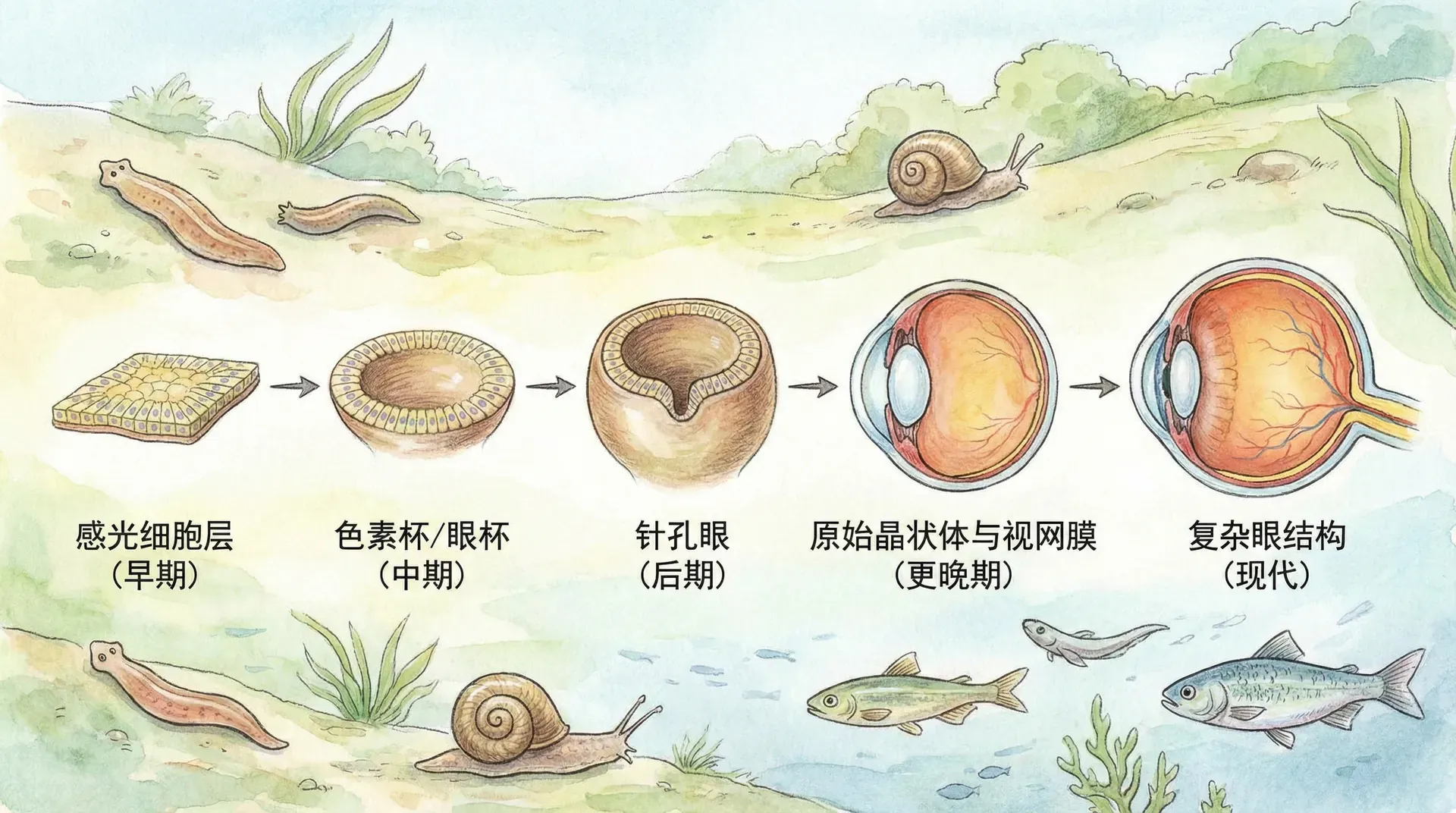
视觉系统的进化历程
部分视觉的价值
关于复杂结构渐进进化的怀疑者最常见的问题之一是:“5%的眼睛有什么用处?”这种质疑实际上忽略了一个关键事实——所有的生物结构和功能,哪怕只完成了一小步的进化,只要能略微提升生物的适应性,都会被自然选择所积极利用。
当你近视忘记戴眼镜,视野虽然模糊,但依然可以辨别物体轮廓、避开障碍物,甚至还能察觉光线变化。你绝不会因此选择闭眼不看——模糊的视觉总比完全失明有用。这一原则其实贯穿所有视觉能力的形成过程。
在自然界中,道理更加直接。对于捕食者和猎物来说,哪怕只具备模糊地“看见光和暗”的能力,也意味着能在关键时刻提前避开危险或者提前发现食物。例如即使只有一种针对光线强弱变化的“光敏点”,动物就可以分辨出白天和黑夜,并选择最安全的时机觅食、休息或交配。而5%的视觉功能,往往足以让生物比完全无视觉的同类占据巨大的生存优势。
这种“哪怕只有一点点感知能力都值得”的原则,并非停留在想象层面,而是被多种实验和野外观察反复证实。例如,科学家曾对果蝇的突变种群进行实验,发现它们哪怕只恢复了部分视觉,遇到天敌时的逃避反应就明显增强。
中国动物的视觉系统多样性
中国地域辽阔,生态多样,孕育了丰富的代表性动物。从最低级的光敏细胞,到复眼、到高度专门化的猫头鹰夜视系统,这些生物阶段就像一张“进化阶梯”,完整展现了视觉系统如何由简到繁演化的全部过程。
这些现生动物之间还存在大量“中间状态”:比如许多闽南的深坑鱼类拥有既不完全简单、也未高度复杂的褶皱视网膜。还有如白鹭等水鸟,在复眼结构与人类单镜眼之间展现独特的优化。
正如图表所显示,视觉系统的任何微小改进都直接转化为生存概率的提升。这种“连续累进的适应优势”是自然选择作用的基础。比如,扇贝眼点能察觉阴影使其在捕食者逼近时及时闭壳,而蜻蜓的高清复眼则能在空中精准猎食。每一级的演化,都对应着一部分生态困境的解决方案。
针孔相机眼的演化
除了上述典型的“生物之眼”,中国海域的动物也展示了视觉进化的多样性。例如乌贼、章鱼具有人类般的透镜单眼,但鹦鹉螺却保留了“针孔相机眼”:
鹦鹉螺的眼睛没有晶状体,而是通过一个小孔让光线进入。孔径越小,成像越清晰但亮度不足;孔径越大虽亮但成像模糊。鹦鹉螺的眼正处在这两者的权衡点。这种古老的眼睛,为科学家揭示了复杂视觉器官的早期雏形。
尽管鹦鹉螺数亿年来一直保留针孔眼,却未像乌贼、鱼类等演化出高度复杂的晶状体眼。这说明生物结构的演化不仅受自然选择驱动,还受限于其独特的遗传与发育方式——发展晶状体眼的“路线”对某些生物可能物理上就难以实现。
鹦鹉螺的“针孔眼”恰恰说明:视觉系统的每一个微小改良,都能带来边际适应性提升,只要在生态位中有用,不一定都要进化到“完美”形态。
其他器官系统的渐进演化

翅膀的起源
“半个翅膀能干什么?”这是进化论反对者喜欢用来挑战渐进理论的问题。但现实中,拥有“半翅膀”的动物极其常见,只是它们的翅膀并非用来飞,而是具备其他生存优势。
以动物为例:
实际上,很多昆虫及无脊椎动物,比如中国的竹节虫、纺织娘等,都有发育不完全的半翼,能帮助短距离游移、逃避天敌。鸟类的雏鸟翅膀也显示出滑翔——扑腾——腾飞的典型渐变过程。
肺部的演化历程
呼吸系统的进化历程堪称渐进优化的典范。最早的脊椎动物只是在消化道附近的一小段囊泡(气室)里储存空气,以应对临时缺氧。但这个结构的“初级版本”只需要很简单的突变就能获得生存优势。
中国内陆水域多缺氧,有趣的各种“初级肺”就在此广泛出现。比如:
- 黄河、长江等浑浊河流的鲤鱼、青鱼,为了对抗水中低氧,会浮到水面吸气,将空气短暂存储在口腔或鳃腔。这类简单“空气室”即为肺的演化起点。
- 原始肺的进一步演化体现在气室内壁布满血管、小突起,有效提升氧气与血液的交换效率。
- 部分鱼类继续优化这一结构,衍生出多室分隔、分支管道,极大提升了氧气利用率。
- 现代鱼鳔,原本用于辅助呼吸,后来被“征用”为调节浮力的新器官;而肺鱼类则延续了“真正肺”型呼吸系统,其后代最终发展为两栖类与爬行动物高度复杂的肺泡结构。
这一渐进过程里,每一小步都只是轻微结构和功能调整,却都能在极端环境下带来明显的生存提升。事实上,鱼类、两栖动物到爬行动物的化石序列,为这一“连续进化”提供了实证支持。
听觉系统的发展
听觉的进化同样是一级级改进的成果。最原始的生物只需要能感觉某种振动就足以获益:
- 蛇类虽然没有典型耳廓,但下巴骨通过头骨直接接触地面,可以灵敏地感知猎物或天敌带来的微振,这种“传震”感受是听觉和触觉的过渡形态。
- 蜘蛛的丝网可放大风和生物带来的振动,蛛丝上的各类感受器细胞可以分辨环境的各种信号类型,如猎物挣扎、风力大小、同类求偶等。
- 春天的蝈蝈和纺织娘等昆虫,通过前足的鼓膜结构“听”到同伴鸣叫,这是一种高级“听觉器”的简形。
- 哺乳动物(如蝙蝠)的听觉极度发达,外耳、内耳、听小骨到大脑声音处理区,无不是一系列微细演化积累之产物。蝙蝠更能发展出回声定位系统,实现三维空间精准导航。
不仅如此,人类胚胎、鱼类、两栖类和爬行动物等耳朵和听觉系统的发育过程,也高度对应了这些渐进演化的“回顾”,被称为“生物发生重演律”。
值得强调的是,哪怕只有5%甚至1%的听觉能力,也远比毫无听觉更具生存竞争力。野外实验甚至表明,听力仅部分受损的老鼠,逃离天敌的反应比听觉完全丧失的个体高出几个数量级。每一处功能的小幅提升,都是自然选择能够“抓住”的机会。
趋同演化
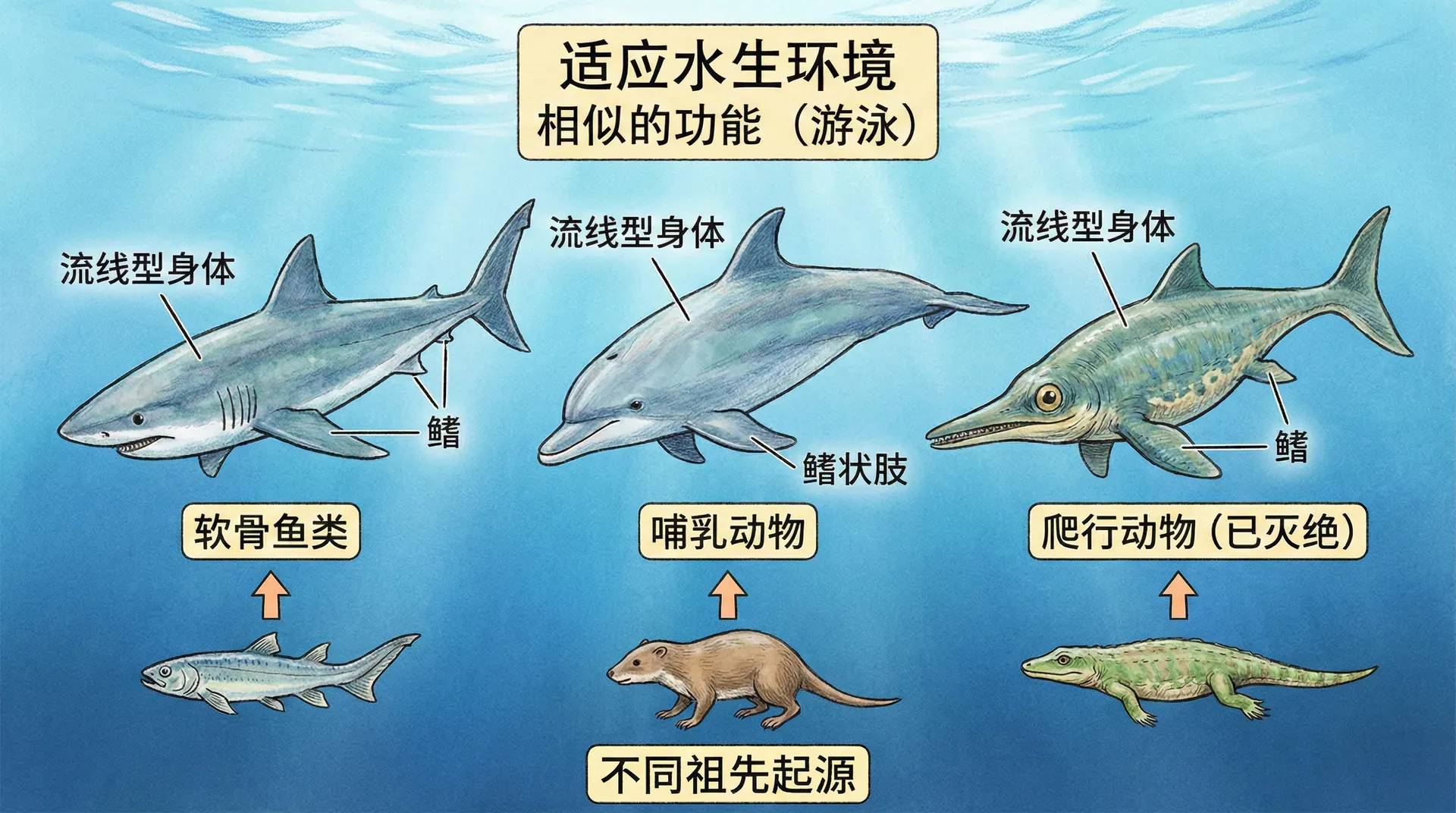
趋同演化指的是不同亲缘关系的生物,在相似环境压力下,独立演化出相似的结构或功能的现象。它展示了自然界面对类似生存挑战时所出现的“解决方案趋同”现象,是理解进化论的重要窗口。“独立创新”往往会在生物界不断上演。
回声定位的独立发明
回声定位无疑是趋同演化中极具代表性的例子。在多个与亲缘无关的动物门类中,这一复杂感官系统都以各自独特的方式被“发明”出来,大大增强了它们在极端环境下的感知能力。
在中国,可以观察到至少两类具有回声定位能力的动物:
1. 中华白海豚
栖息于珠江口等浑浊水域,水下能见度极低。传统视觉根本无法定位猎物。
进化出发达的回声定位系统,包括:
- 精密的发声结构(额部的“声波聚焦器”):可发出高频超声
- 特殊的接收器官(下颌里的脂肪垫,能导声至内耳)
- 神经处理系统:大脑相关区域极度发达,可将回声讯号还原为空间图像
海豚依靠回声定位实现高精度导航、觅食和族群交流。甚至能通过回声分辨出鱼群密度、个体形状和大小。
2. 食虫蝙蝠
生境类型多样,包括山洞、丛林乃至城市。夜间活动,光线极弱。
进化出高度发达的回声定位能力,但结构与海豚完全不同:
- 发声方式:通过喉部(而非头额部)发出超声波,经口鼻喷出
- 接收方式:外耳巨大、灵活,能“追踪”声波;下颌骨参与导声,但机制不同
- 大脑处理机制:听觉区域专门化,适应信号处理,与海豚具有本质差异
蝙蝠依靠回声定位避障、捕猎飞虫,有的种类还能检测到微小昆虫的翅膀振动。
对比分析:
更广泛地说,鸟类中的雨燕、金枪鱼等部分物种也独立演化出原始的声呐机制,用于特殊环境下定位。回声定位的多次“独立发明”,充分体现了自然选择对有效感知手段的“高效收敛”。
如果某种设计极为有效,自然界就会在不同进化分支中反复地“独立发现”它。
电场感知的趋同演化
类似地,不同动物还独立进化出“电场感知”这种几乎不可见的第六感。
1. 长江中华鲟
- 生活在水流湍急、泥沙众多的长江水域,依赖电感受器协助发现猎物。
- 感知机制:主要是皮肤头部和体侧特殊细胞,被动感知水中微弱电场变化。
- 主要功能:定位猎物、导航迁徙,在能见度极差时依然能精准行动。
2. 南美亚马逊的刀鱼和电鳐(如电鳗)
- 独立进化出强大的电生理系统。有些鱼类甚至能主动产生电场,通过检测自发电场的扰动主动感知世界。
- 感知器官:全身散布电感受器,部分可主动发电,部分被动物用于捕食或防御
- 脑部处理:专门神经区域强化处理电信号
- 功能相似:精准觅食、避障和水域导航
趋同演化不仅发生于感官系统。比如大熊猫的“伪拇指”与鼩鼱(袋熊等)的类似结构、沙漠中不同植物门类的多肉化、狼与袋狼的形态趋同等,都是自然选择多次“独立走通相似路径”的例证。
渐进演化的时间尺度
理解演化的渐进性,必须结合“地质时间”的广阔尺度。人类微观的时间观让我们难以想象生命可以在数百万年间由简单逐步走向复杂。
如图所示,每一代生物的微小优势都会在自然选择下累积。尽管单次改进也许微不足道,但成千上万代后,这种细小“步进”可酿成巨大飞跃。
例如,哺乳动物的中耳小骨就是由早期爬行动物下颌骨结构逐步细化、演化而来,并非一蹴而就。植物的叶片气孔分布、树木维管系统、昆虫的复杂翅膀、甚至人眼的高度精巧,也都见证了“细微变异+巨量时间”所能达成的奇迹。
自然选择之所以强大,正因为它擅长“放大”每一代微小的、可遗传的有益变化——不断积累下去,最终让结构和功能复杂度出现质的跃升。在漫长的地质年代里,几乎没有不可能的进化“突破”。
理解演化的原理
综合上文,我们可以归纳出现代生物学解释复杂结构起源的几个核心原理:
这些原理共同构成了现代进化论的内核。无论是大熊猫手掌的“假拇指”、中华白海豚的声纳、红嘴鸥的候鸟导航、还是金丝猴的高原气血调节,全部都是细微累积与选择积淀的成果,并非某种瞬时出现的“设计智慧”。
理解渐进演化和趋同演化,不仅能帮助我们欣赏每一个生物的独特美,更让我们明白:生命的复杂和精巧是自然过程耐心雕琢、无数代选择与时间的产物。哪怕没有智能“工程师”,自然选择和随机变异也足以孕育出远超人类想象的生命奇迹。