叶绿体基因表达系统
叶绿体是植物细胞中最为核心和独特的细胞器之一,被誉为植物生命活动的动力工厂。其最显著的功能是在光合作用过程中,将太阳光能高效地转化为有机化学能,为植物的生长和发育提供能量基础。通过这一过程,叶绿体不仅推动了植物体内碳的同化,更通过合成淀粉等储能物质,为粮食作物的产量和品质打下坚实基础。
在我国广袤的农田,南至江南水乡的水稻,北至黑龙江玉米带,西至黄土高原的小麦,每一片绿色叶片中都蕴含着数以亿计的叶绿体,它们默默地将阳光“收割”为人类赖以生存的食物。叶绿体普遍分布于叶肉细胞、幼枝以及发芽的幼苗之中,它们的活跃与否直接决定了作物的生长速度与产量高低。
叶绿体不仅是光合作用的中心场所,还承担着诸如氨基酸合成、脂肪酸代谢、色素与激素合成、硫和氮的同化等多项复杂生理和代谢活动。正因如此,叶绿体常被称作植物细胞中的“生化工厂”,其多功能属性使其对植物整体生理状态和抗逆性具有根本影响。
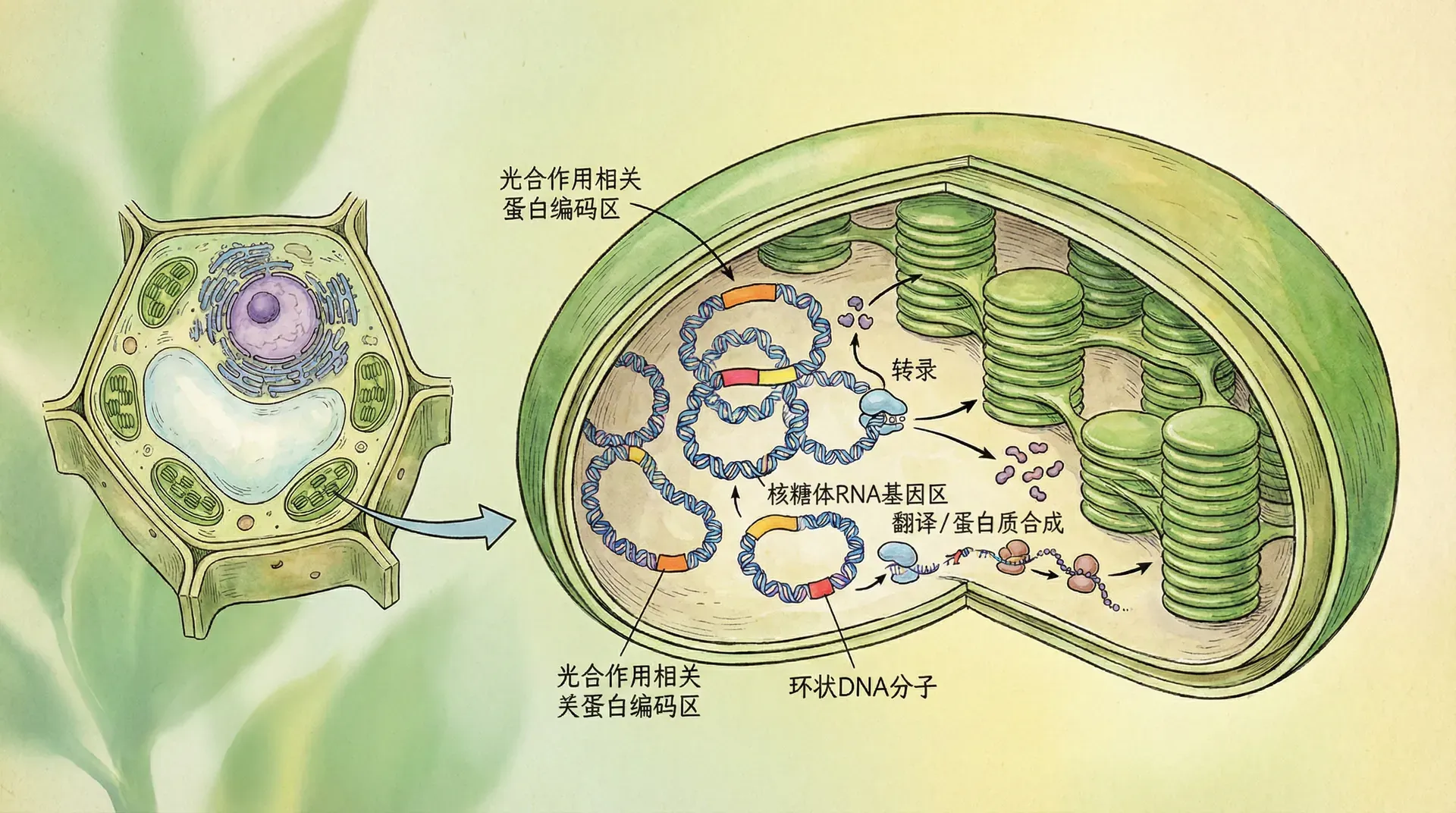
从结构上来看,叶绿体具有独特的双层膜结构,将其与细胞其它部分隔离开来。内部富含层状的类囊体系统,有助于高效吸收和转化光能。更重要的是,叶绿体拥有独立的遗传物质——自身的基因组。尽管叶绿体基因组较小(一般约为120-160 kb,总体不超过150 kb),但却保存着大约100个功能基因,主要负责编码光合作用需要的蛋白以及与转录、翻译、代谢密切相关的分子。与细菌基因组相比,叶绿体在进化过程中经历了大规模基因丢失和转移,但也逐步发展出复杂且高效的遗传调控机制,实现了植物高等化的适应之道。
在数量分布上,叶绿体是植物细胞中最丰富的细胞器之一。例如水稻,一个叶绿体通常含有约100份基因组拷贝,而每个叶肉细胞中可包含几十至逾百个叶绿体,总计算下来单个细胞中的叶绿体基因组拷贝可轻松超过1万份。这种富集的遗传资源,为高强度的光能转化及蛋白合成提供了充足物质支撑,也为植物对逆境的快速响应和适应积累了潜力。
值得注意的是,不同作物的叶绿体基因组在结构和功能基因布局上进行着微妙的差异化进化,以适应东西南北不同的气候和生态环境。例如,玉米叶绿体基因组略大于水稻,且蛋白编码基因数略有增加,反映出C4作物光合作用效率更高的分子基础。
双重转录系统的协调工作
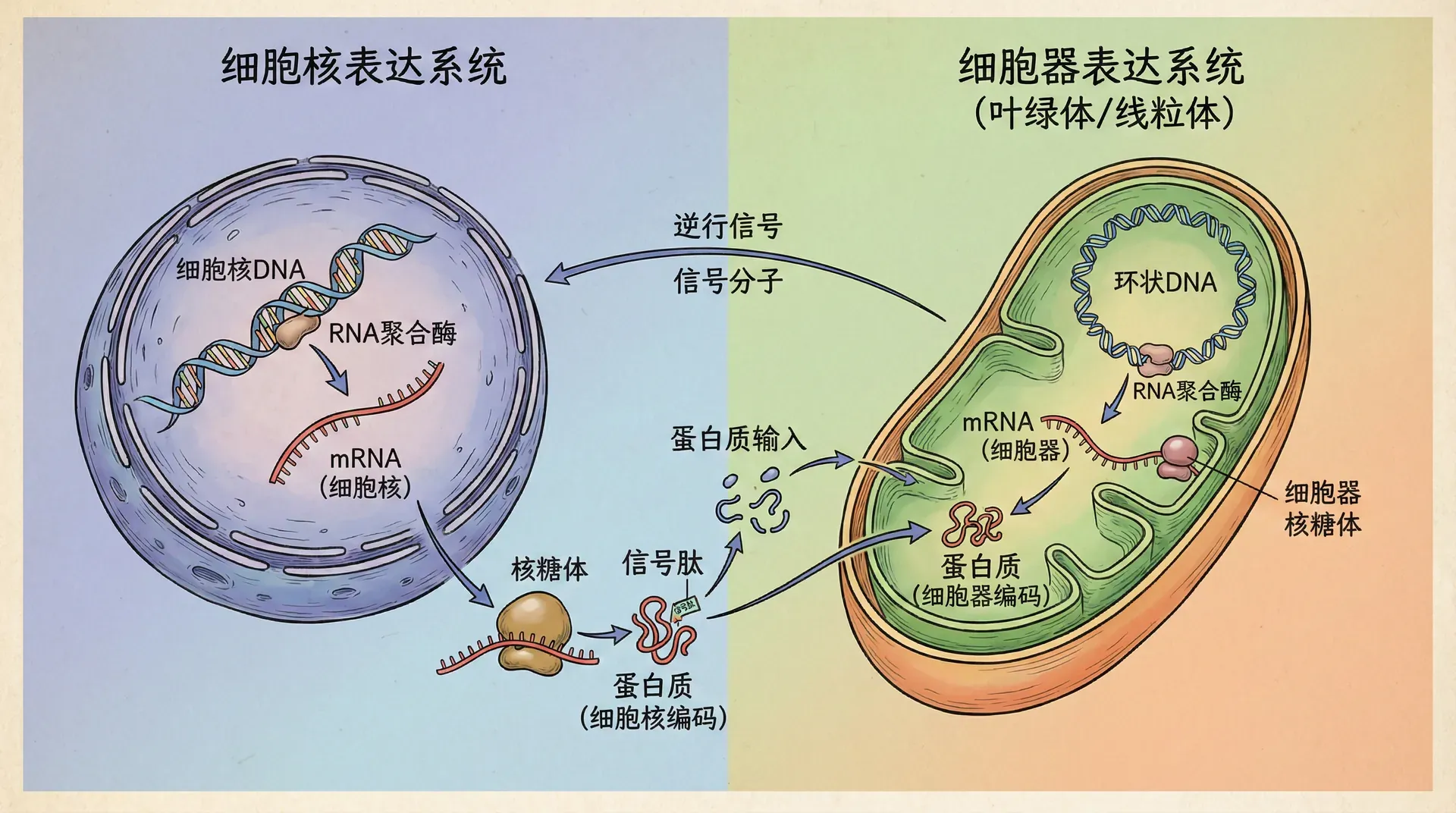
叶绿体基因表达的最大特色是同时具备了两套结构和起源完全不同的RNA聚合酶系统,并协调运作。这两套系统,即质体编码RNA聚合酶(PEP)与细胞核编码RNA聚合酶(NEP),共同维持着叶绿体在各发育阶段及复杂环境条件下的生理平衡和基因表达弹性。
质体编码RNA聚合酶(PEP)主要负责那些与光合作用直接相关的基因的转录,包括反应中心蛋白、光系统I、II等;而细胞核编码的RNA聚合酶(NEP)则偏重维持细胞基础功能和结构相关的“管家型”基因表达。
在叶绿体的发育历程中,如以水稻为例,PEP和NEP的活性并非静态不变,而是经历着动态的时序调控。幼苗期由于光合作用尚未完全建立,NEP相对活跃,优先合成叶绿体结构和基本维持所需蛋白,为后续光合作用功能的建立奠基。待到植株进入快速生长期,叶片功能趋于完善,PEP活性迅速上升,主导光合作用蛋白的大量表达,以满足能量转换和同化物合成的剧增需求。
这种交替主导和动态调控,不仅是植物生长发育的需求使然,更是对外部环境变化的一种适应。以水稻为例,在苗期种子内贮存的养分尚能满足新生组织的基本需要;但随发育推进,外部光能成为植物生存的决定性条件,PEP系统的激活直接关联到光合速率的提升。
RNA聚合酶系统的分子机制
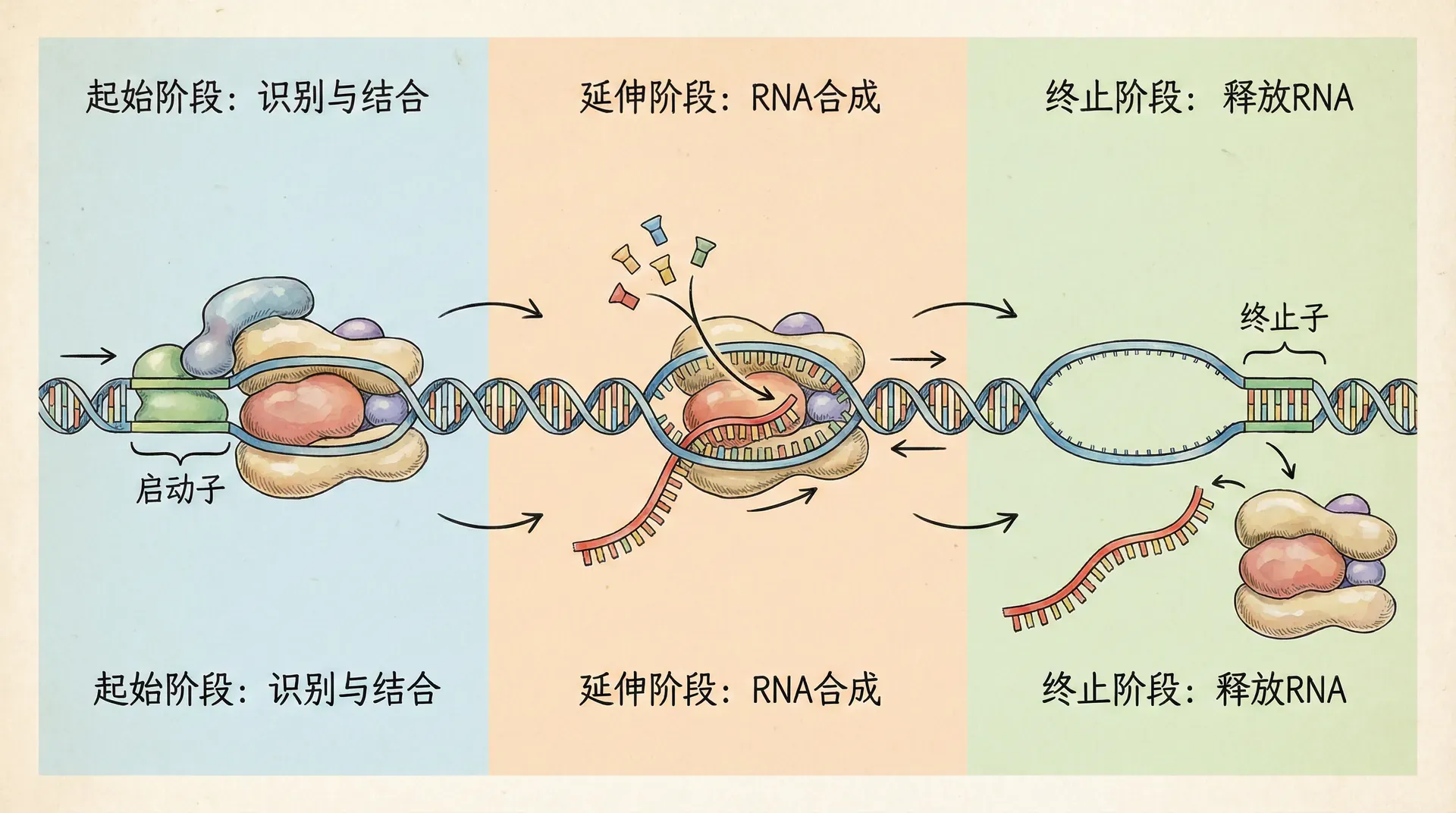
PEP和NEP系统在分子结构与调控方式上各具特点。PEP系统与蓝藻和细菌的DNA依赖型RNA聚合酶同源,由多种亚基组装而成,并在高等植物中进一步集成了十几种核编码的辅助蛋白,如PAP(PEP-associated proteins)。这些辅助蛋白增强了PEP对复杂启动子的识别与调控能力,并使PEP成为一个重量超过1兆道尔顿(MDa)的巨大转录复合体。成熟叶片中,PEP不但负责光合相关蛋白的高效表达,还对一系列逆境应答基因具有调节功能。
NEP系统则结构更趋简单,是单亚基的RNA聚合酶,功能上类似于噬菌体T7 RNA聚合酶。因其分子量较小、结构灵活,NEP能迅速感知和响应外界信号变化,及时调整基础代谢与叶绿体复制过程中的必需基因表达。两者之间通过极为复杂却高效的信号网络实现分工协作,显著提升了叶绿体的基因表达多样性与调控精细度。
叶绿体发育中的基因表达模式
叶绿体基因表达的时空变化,在不同作物与发育阶段表现出极具规律性的分布。以小麦为例,从种子萌发到绿叶完全展开,基因表达的高低随功能定位和外部刺激不断调整。早期阶段,维持叶绿体结构、复制及转录翻译的“管家基因”如rpoB、clpP高水平表达,保证基础功能的建设和维持;光合作用相关基因如psaA、psbA则于叶绿体成熟过程中逐步上调,到叶片伸展、光合功能需求顶峰时达最大值。
这些表达模式不只是动态数据的变化,更蕴含着叶绿体进化和功能分化的精妙机制。稳态的管家基因表达确保了基础代谢与细胞器结构的健全,而光合作用相关基因的时空激活,则保证了植物在合适时间点具备最优光合性能。这种分级、分区的精细化表达,使得植物能适应多变环境和不同生长速率的需求。
光环境对基因表达的调控
光照不仅决定了植物形态,也精准塑造了叶绿体基因的表达谱型。不同波长、强度与周期的光照会激活特定的信号转导通路,改变PEP和NEP两套系统的相对权重,最终体现在光合能力的增强或抑制。
系统研究发现,蓝光信号主要通过隐花色素(cryptochrome)受体影响叶绿体的整体转录活性,推动相关基因高峰表达;而红光则依赖光敏色素(phytochrome)系统,对某些特异性光合和生理调控基因进行精细调节,从而实现对环境的最优适应。
这种光质调控模式,不仅对野外复杂气候下的作物至关重要,在现代农业特别是在设施农业(如温室种植)中同样发挥巨大作用。通过调控人工光源(如LED蓝、红光比例),农艺师能够有针对性地诱导叶绿体表达最有利于产量和品质的基因,实现精细化管理与高效生产。例如南方夏季强光下,叶绿体主动上调光抑制保护蛋白的合成,以防细胞受损;而北方冬季弱光环境中,则促进光能捕获及能量转化相关蛋白的表达。
叶绿体基因表达的生物学意义
叶绿体独特的“双重转录系统”与复杂灵活的表达网络,不仅保证了植物在正常生长发育中的光合效率和能量转换能力,更为作物因应各种逆境(如强光、低温、干旱、病虫害)提供了强大分子基础。叶绿体基因表达的动态调控,是提高农作物产量、改善品质和拓展种植区域的关键分子底盘。
近年来,随着转录组测序、蛋白质组学及分子生物学技术的飞速发展,我们对于叶绿体基因表达调控已从宏观到微观、从时空到网络结构有了更系统和定量的认知。在全球气候变暖、耕地资源受限的背景下,深入研究和精准调控叶绿体的表达途径,必将助力于高产抗逆农作物的选育,同时也为生物能源开发、环境可持续管理等领域开辟新前景。
实际应用与展望
随着分子生物学、转录组学和基因编辑等前沿技术的不断进步,叶绿体基因表达调控的研究已成为作物分子育种和高效农业的重要突破口。近年来,科学家们在水稻、小麦和玉米等主要粮食作物中,通过靶向调控叶绿体基因表达,取得了一系列具有实际应用价值的成果。
首先,通过基因工程途径调节叶绿体内关键转录因子或调控蛋白(如PEP相关蛋白、NEP因子等)的表达水平,显著提高了植物的光合效率。例如,在水稻中过表达PAP(PEP-associated protein)不仅增加了叶片的光合作用能力,还提高了整体产量和对逆境的耐受性。类似策略还被用于小麦、番茄等作物,有效提升了在弱光、盐碱、干旱等逆境环境下的生长表现,这为发展适应密植和复杂气候的新品种提供了理论和技术基础。
此外,叶绿体基因工程还被应用于营养品质改善。例如,通过精准调控叶绿体基因通路,增强类胡萝卜素、维生素等营养因子的合成与积累,未来可望培育出更富有营养价值的作物。此外,有研究在叶绿体中高效表达新型蛋白或工业酶,推动了植物工厂和绿色生物制造的发展,为生物能源和新材料领域拓展了思路。
然而,叶绿体基因表达的调控网络极其复杂,任何人为干预都需要经过严格的安全性评估,确保不会对植物的其他生理功能产生负面影响。不同作物、不同发育阶段对调控机制的敏感性也存在较大差异,未来需要加强基础研究,以规避潜在的生态和食品安全风险。
展望未来,叶绿体基因表达调控的研究与应用有望在以下几个方向持续发力和深化:
除了提升产量和品质、增强环境适应性之外,叶绿体作为碳固定的核心场所,在全球气候变化背景下还有助于碳中和和生态环境保护。未来,结合人工智能、大数据分析与系统生物学,开发可以实时感知环境变化并动态调控基因表达的“智能叶绿体”系统,将成为新一代智慧农业和作物分子设计育种的核心目标。
通过对叶绿体基因表达调控机制的深入挖掘与精准操控,我们不仅能够培育出高产、优质、资源利用高效的农作物,还有望推动绿色能源、生物制造等领域的技术革命,为保障人类粮食安全、实现可持续发展和应对全球性能源危机提供坚实的分子基础和广阔的发展前景。